Оглавление Введение Часть 1 Часть 2 Часть 3 Заключение
Глава 1 Глава 2 Глава 3 Глава 4 Глава 5 Глава 6 Глава 7 Литература
Глава 4
Гомеостаз клетки
Клетка[1] является основной структурно-функциональной единицей всех живых организмов и определяется как «элементарная» живая система[2]. Размеры клеток варьируют от 0,1 мкм (10-3 см) до 155 мм (1,55×101см). Клетка может существовать и как отдельный организм[3], так и в составе тканей многоклеточных животных, растений, грибов. Лишь вирусы представляют собой некоторые неклеточные формы «жизни».
Содержимое клетки – цитоплазма, содержащая генетический аппарат, как носитель наследственной информации, который в клетках эукариот[4] заключен в ядре, отделенном сложным мембранным аппаратом от цитоплазмы. В клетках прокариот[5], лишенных оформленного ядра, генетический инструмент расположен в нуклеотиде. Клетки эукариот способны к самовоспроизведению путем митоза[6], а половые клетки образуются в результате мейоза[7].
К характерным и уникальным особенностям, присущим всем клеткам, следует отнести: пространственную организацию биохимических процессов, их компартментализацию[8], микрогетерогенность и компактность. Эти свойства обеспечивают высочайшую эффективность, упорядоченное расположение, строго последовательное сопряжение и, одновременно, разделение различных биохимических процессов в миниатюрном общем объеме цитоплазмы при постоянном использовании одних и тех же элементарных предшественников в общем динамическом процессе ассимиляции, диссимиляции, обмена и транспорта веществ[9], энергии и информации.
Благодаря такой активной, гибкой и скоординированной деятельности внутриклеточных структур и образований, демонстрирующих высшую структурно-функциональную форму систематизированного неравновесия, осуществляются и непрерывно поддерживаются значительные ионные градиенты внутри и вне клетки, создающие беспрецедентные условия для формирования трансмембранных внутриклеточных потенциалов, колеблющихся от 2-5 мВ до 140-200 мВ, демонстрируя таким образом, историческую целесообразность энергообразующих и реактивных статических сил в сложнейшем гомеостатическом устройстве живой материи.
Все клетки эукариот имеют сходный набор органоидов и органелл, сходно регулируют метаболизм, запасают и расходуют энергию, сходно с прокариотами используют генетический код для синтеза белков и собственного воспроизводства. У эукариотных и прокариотных клеток принципиально сходно функционируют и клеточные мембраны, наделенные способностью всех видов трансмембранного транспорта: от банальной и облегченной диффузии, пиноцитоза и фагоцитоза до активного транспорта органических соединений и ионов против градиентов их концентраций.
Общие признаки клеток свидетельствуют о единстве их происхождения, хотя могут очень отличаться по своим размерам, форме, числу тех или иных включений, наборов ферментов и т.д. Это обусловлено последствиями кооперирования клеток в многоклеточном организме и, соответственно, с одной стороны, дифференцированием их функциональных обязанностей, т.е. специализацией функции, с другой, как результат такой специализации[10], существенные морфологические отличия[11].
Различия в структуре и функциях одноклеточных организмов также в значительной степени связаны с их долговременными приспособлениями к среде обитания, но без учета адаптивных возможностей и преимуществ кооперации[12].
Довод в пользу единого происхождения прокариотических и эукариотических клеток одно и многоклеточных организмов заключен в принципиальном сходстве их генетического аппарата, хотя у различных одноклеточных могли быть и разные прокариотные родители. Согласно гипотезе симбиогенеза[13], одни прокариоты, внедрившись в эукариотные клетки на относительно раннем этапе эволюции, преобразовались внутри клеток хозяина в митохондрии, другие – в хлоропласты и стали самовоспроизводиться как органоиды.
Это далеко не единственная версия в трактовке развития исторических событий. Рассматривается даже гипотеза о постепенном преобразовании собственных структур прокариотной клетки в процессе её превращения в эукариотную[14].
Регулирующие факторы внутри клеток – метаболиты, ионы, продукты активной скоординированной деятельности внутриклеточных органоидов и органелл, которые действуют на гены, приводя к изменению количества ферментов, или на сами ферменты, меняя их активность. Т.е. совершенно очевидно, что и на этом уровне организации материи регуляция гомеостатических реакций осуществляется по принципу обратной связи, когда продукт реакции определяет её интенсивность (скорость реакции) через константу равновесия или константность диссоциации (через норму реакции). В результате такой саморегуляции поддерживается оптимальный уровень многих жизненно важных внутриклеточных процессов, иногда даже при значительных изменениях во внеклеточной среде.
Регулирующие же факторы гуморальной, нервной, нейрогуморальной регуляции деятельности организма[15], действующие на клетки, а также влияние клеток на клетки[16] в пределах прямых или опосредованных контактов составляют базовую основу их возможностей и внешних резервов, поскольку в условиях изолированной культуры, клетки очень скоро[17] утрачивают многие черты своих специализаций[18].
Клеточный гомеостаз высших животных и человека на примере кардиомиоцита[19]
С точки зрения нормальной и патологической физиологии, система поддержания клеточного гомеостаза, в частности высших животных и человека, складывается из пяти основных уровней регуляции:
1. Нервной, гуморальной, объединяемой понятием нейрогуморальная или центральная регуляция
2. системной
3. органной
4. тканевой и
5. собственно внутриклеточной (рисунок 23).
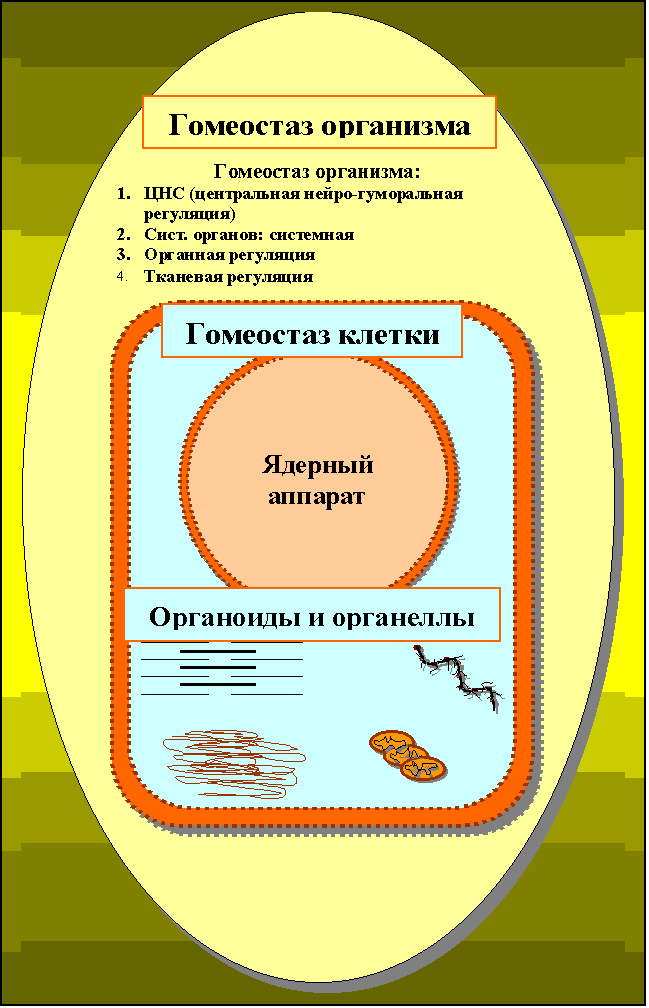
Рисунок 23. Иерархическое устройство отношений гомеостатической системы: клетка – организм, где гомеостаз клетки окружён гомеостазом организма.
Именно они обеспечивают взаимодействие двух интересующих нас носителей гомеостаза: гомеостаза клетки и гомеостаза организма. Причем взаимодействие названных уровней такой регуляции обеспечивает поддержание только одного, конечного и, в то же время, ключевого звена во всей цепи сложных регуляторных отношений: поддержания стабильности внутри- клеточной среды (внутриклеточного гомеостаза), обеспечивающей максимальную защищенность и скоординированность деятельности всех метаболических преобразований, совершаемых во внутриклеточных органоидах и цитоплазме.
В результате взаимодействия этих систем формируется мобильная система реакций фенотипической адаптации[20], объединяющая все уровни гомеостатической регуляции и заключенная в рамки другой, более инертной – генотипической20,[21], обеспеченной информацией, содержащейся в ядерной ДНК. Именно эти, подкрепленные «структурной основой»[22], процессы, получили в курсе нормальной и патологической физиологии свою классификацию: фенотипическая и генотипическая адаптация21,22,[23].
С позиций нашего изложения этот факт следует рассматривать как главный и определяющий доминирующую роль клеточного субъекта в отношениях двух рассматриваемых гомеостазов: первичного (клетка) и вторичного (организм), составляющих по сути, единую высокоорганизованную композиционную или композитную систему.
Для успешной реализации своих функциональных задач клетка использует очень совершенную структуру, выполняющую сложную коммутативную роль, - передачу и проведение информации, осуществляя строго сбалансированный обмен веществом и энергией[24],[25],[26]) с внешней для неё средой. Это плазматическая мембрана или плазмолемма. Именно она обеспечивает непрерывный и контролируемый обмен веществом, энергией и информацией с внешней для неё средой - внеклеточным (межуточным) пространством. При этом, высокая подвижность[27],[28],[29],[30] и непрерывное обновление высокоорганизованной мембранной структуры20,[31],[32], делает этот обмен динамичным, гибким, вариабильным.
Несмотря на то, что системы регуляции более высокого уровня организации (тканевая, органная, системная и нейрогуморальная) выполняют самый сложный комплекс задач, связанных с координацией деятельности всех систем, органов и тканей в организме, а также обеспечивают адекватный обмен энергией, веществом и информацией самого организма с внешней средой, находящейся на ещё более высоком иерархическом уровне - с экосистемой планетарной биосферы, их роль в конечном итоге замыкается всё-таки на поддержании клеточного гомеостаза. Это их главная цель и главное условие их существования. Иными словами: поддержание клеточного гомеостаза есть главное условие существования всего организма в целом.
В таком контексте роль систем регуляции более высокого иерархического уровня ограничивается скромной (вспомогательной) функцией мобильного дополнения к системе внутриклеточной регуляции, как внешнего источника вещества, энергии и информации, стабилизированного гомеостазом всего организма и гомеостатическим поведением особи. Такое построение систем регуляции гомеостаза клетки обеспечивает очень высокую защищенность внутриклеточной среды от неблагоприятных внешних воздействий, иллюстрируя высшую форму своего развития в эволюции адаптивно-приспособительных реакций[33].
Таким образом, если рассматривать регуляторные системы организма в целом с позиций физиологии и клеточной теории, их реальной регуляторной единицей является клетка, реальным объектом регуляции - внутриклеточная среда, реальным органом исполнения регуляторной функции - плазматическая мембрана, а реальным регулятором внутриклеточного гомеостаза - ДНК.
С другой стороны, резервные и управляющие возможности внутриклеточной регуляции значительно расширены мощными регуляторными и «структурно-материальными»15 возможностями систем, принадлежащих к более высокому иерархическому уровню организации материи, составляющей целостный организм. Это системная, органная и тканевая регуляции, представляющие независимый гомеостаз организма, генетически детерминированного от органоидов и органелл до уровня вышестоящих иерархов, действующих как извне (через внешнюю среду), так и изнутри (через отношения в элементарном мире).
Это тем более важно, что регуляторные возможности клеток, попавших в экстремальные ситуации, какими можно назвать наиболее часто встречающиеся для клетки: тотальную ишемию, гипоксию, искусственно вызванную деполяризацию, действие холода, состояния инициальной или прогрессирующей гиперфункции[34],[35],[36] и т.д., чаще всего не могут быть реализованы в полном объеме. Т.е. состояния, связанные с дефицитом кислорода (энергии), вещества (субстрата) и информации (денервация, тотальная ишемия, изоляция) существенно ограничивают их регуляторные и адаптивные возможности.
Рассмотрим некоторые из таких состояний более подробно на примере кардиомиоцита, как наиболее благоприятной модели для анализа.
Пример 1. Состояние тотальной ишемии миокарда, с которым чаще всего приходится сталкиваться в кардиохирургии при выключении сердца из системы общего кровообращения, приводит к фактической редукции регуляторных систем до уровня клетки, где могут быть раскрыты возможности только системы внутриклеточной регуляции. Это состояние можно охарактеризовать как одно из наиболее тяжелых в смысле резервных возможностей. В этом случае процессы поддержания стабильности внутренней среды (гомеостаза) вынуждены протекать в условиях очень жестоких: в условиях абсолютного дефицита внешних поступлений вещества, энергии, и, фактически, информации.
Пример 2. Состояние искусственно вызванной кардиоплегии, наиболее широко используемого сегодня метода противоишемической защиты сердца во время операций, можно охарактеризовать почти также. Разница лишь в том, что в этих условиях более глубоко и более резко наступает деполяризация мембран кардиомиоцитов[37],[38], а это сопряжено с опасностью потери клетками одной из форм «свободной» энергии - энергии трансмембранного потенциала21,24,[39]. Как показали наши эксперименты, деполяризуемая клетка в борьбе за сохранность своего трансмембранного потенциала на каком-то предельном уровне немедленно прибегает к усиленному расходованию энергии АТФ на его частичное восстановление29,30. Удалось даже определить этот «критический уровень» поляризации –20 – 40 мВ.
Пример 3. Состояние гипоксии, всегда сопровождающее кардиологические патологии, наверное, целесообразно рассматривать исходя из наиболее известной и принятой классификации16,[40], поскольку в их этиологии лежат различные механизмы, по-разному затрагивающие рассматриваемые нами процессы регуляции клеточного гомеостаза.
Пример 4. Состояние цитотоксической гипоксии наиболее губительно для систем регуляции, поскольку сопряжено с ограниченной возможностью утилизировать энергию, субстраты и адекватно воспринять поступающую информацию. Это происходит за счет нарушений, которые принято называть «метаболическими»16. Но, главное в том, что в случае цитотоксической гипоксии поврежденными (или блокированными) оказываются практически те же реакции, которые обеспечивают поддержание стабильности внутренней среды клеток (те самые «метаболические реакции» , о которых упоминалось выше). Таким образом, страдает собственно система внутриклеточной регуляции гомеостаза.
Пример 5. Бактериальная интоксикация. Так при дифтерийной или химической (2,4-динитрофенолом) интоксикации были отмечены резкие, хотя и разной интенсивности, морфо-функциональные нарушения27. Но в том и другом случаях эти изменения затрагивали сферу гомеостатических реакций. Так дифтерийная интоксикация сопровождалась сначала умеренной активацией сукцинатдегидрогеназы (СДГ) и глюкозо-6-фосфодегидрогеназы (Г-6-ФДГ) рабочего миокарда левого желудочка сердца.
Это типическая реакция, вызванная гипоксией и связанная с попыткой ее компенсации за счет активации «быстрого» участка в цикле Кребса[41] и шунтирующего сброса протонов на ФАД-зависимые системы[42], а также за счет естественной в данной ситуации активации гликолиза и гликогенолиза22. В последующем активность СДГ и Г-6-ФДГ несколько падала, что это можно было бы связать с компенсацией дефицита энергии, но этого не происходило[43].
При химической интоксикации первичное повышение активности всех ферментов, включая СДГ и Г-6-ФДГ, было незначительным и неравномерным, а затем сопровождалось резким падением, особенно СДГ. Причем все эти колебания активности происходили также на фоне нарастающего дефицита энергии и нарастающей гиперфункции35. Это уже серьезный признак развивающейся декомпенсации сердечной деятельности. Так и было: эффективность функционирования структур резко падала, развивалась сердечная недостаточность кровообращения и на 7-10 сутки от начала эксперимента животные погибали35. Это указывает, прежде всего, на уменьшенную мощность регуляторных систем, ослабленную их цитотоксическим повреждением.
Пример 6. Цитотоксическая гипоксия. В этиологии цитотоксической гипоксии возможны также и случаи блокады (или повреждения) рецепторных зон плазмолеммы. Тогда нарушаются процессы восприятия информации, и система клеточной гомеостатической регуляции теряет возможность адекватно реагировать на внешние раздражения, лишаясь связи в получении управляющей информации от регуляторных систем более высокого уровня организации. Примером этому в рассматриваемых нами условиях, могут быть ситуации, связанные с искусственной блокадой альфа- и бета-адренорецепторных зон аденилатциклазы[44],[45] фармакологическими средствами, в частности, при интраоперационной защите миокарда.
Пример 7. Блокада рецепторных зон. Опыт показал, что введение в состав кардиоплегических растворов бетта-блокатора обзидана (пропранолола) не вызвало сколько-нибудь выраженного усиления противоишемической устойчивости консервируемых сердец[46],[47],[48],[49], поэтому и не нашло потом широкого распространения. Исходно авторы ожидали противоишемического действия бетта-блокаторов за счет уменьшения реактивности регуляторных клеточных систем. Они его и получили, но в очень слабо выраженной форме. С точки же зрения теории адаптации нельзя было и ждать от систем клеточной регуляции существенных изменений в их устойчивости к ишемии. Кардиоплегические растворы, деполяризовав клеточные мембраны, уже почти полностью лишили кардиомиоциты возможности использовать резерв систем клеточной регуляции гомеостазом более высокого уровня организации, поскольку, искусственно редуцировав всю эту систему до клеточного уровня, лишили клетки внешнего источника поступления информации, вещества и энергии. Именно потому дополнительная блокада только одной из структур, участвующих в передаче только части общей информации, теряет всякий смысл. Введение же бетта-блокаторов за 1-2 часа до ишемии, в качестве предварительной фармакологической подготовки к предстоящей ишемии, когда сохранены все структурно-функциональные взаимоотношения систем регуляции клеточного гомеостаза, представляется нам более интересным.
По данным института хирургии РАМН им.А.В. Вишневского[50],[51] и Института сердечно-сосудистой хирургии РАМН им. А.Н. Бакулева[52],[53],[54] предварительное введение бетта-блокатора обзидана (пропранолола) имело определенное противоишемическое действие: такие сердца легче переносили тотальную тепловую ишемию, лучше сохраняли свои функциональные характеристики в восстановительный период. Однако, по более поздним данным, полученным в НИИ трансплантологии и искусственных органов МЗ РФ, основанном на анализе динамики физиологических, биофизических и биохимических характеристик клеточного гомеостаза кардиомиоцитов при таком использовании обзидана, было установлено, что эффект его действия, пролонгирующего процесс умирания сердца, весьма незначителен, выражен в большей степени без сочетания с кардиоплегией и связан, по всей видимости, с активацией шунтирующих процессов в передаче информации (по типу легкой формы гипоксической и циркуляторной гипоксии – «легкий тренинг»).
В интересующем нас аспекте, этот пример может быть прекрасной иллюстрацией эффекта, связанного с блокадой систем внутриклеточной регуляции.
Так большинство авторов, вводивших обзидан в доишемический период в дозах, оказывающих защитное противоишемическое действие (0,05-0,2 мг/кг массы), указывают на резко выраженный его гипотензивный эффект. Это происходит за счет отрицательной реакции хроноинотропии, отражающей состояние адаптивных систем в реакциях Франка-Старлинга и гетерометрической регуляции[55]
Пример 8. Функциональное разобщение клеток: фармакологическая кардиоплегия. В общем контексте нашего изложения, следует сказать, что введение в составы кардиоплегических растворов препаратов мембранотропного типа действия вообще оказывается неэффективным30, хотя несколько лет назад использование таких препаратов как новокаин, лидокаин, прокаин, ксилокаин, хинин и хинидин, нифедипин, вводимых в составы растворов, было весьма популярным. Исключением здесь являются лишь растворы, составленные на основе перфторорганических эмульсий[56], и фосфорилированные метаболиты, в частности, фосфокреатин (или неотон)[57], на действии которых в настоящей работе мы останавливаться не будем). Эмпирические же попытки улучшить результаты интраоперационной защиты миокарда (кардиоплегии) такими средствами, как введение в состав растворов препаратов мембранотропного типа действия оказались не достаточно эффективными и потому не нашли сегодня широкого применения. Многие же медицинские центры, использовавшие новокаин, прокаин, нифедипин, хинин и хинидин для введения в раствор, уже исключили их из своей постоянной практики.
Иллюстрацией к потере клетками способности адекватно реагировать на поступающую извне информацию при использовании таких добавок в кардиоплегические растворы может быть единодушное мнение всех упоминаемых выше авторов о снижении или полном устранении эффекта вазоконстрикции в ответ на введение деполяризующих и охлажденных растворов в коронарное русло. Это и было собственно главной побудительной причиной для добавления указанных препаратов в составы кардиоплегических растворов.
В качестве дополнительной ремарки к вышеизложенному, следует особо подчеркнуть, что использование фармакологических добавок в кардиоплегические растворы с целью пролонгации времени эффективной защиты сердца от тотальной ишемии не является панацеей. И, если авторы, получившие положительный эффект от того или иного вещества, добавленного в раствор, называют его действие эффективным, то в абсолютном выражении эффективность эта не очень значительна.
В своих ранних исследованиях нам также пришлось заниматься эмпирическим испытанием фармакологических добавок в составы кардиоплегических растворов. Тогда нами было испытано более пятидесяти препаратов самых различных фармакологических групп. Результаты были поразительны. Нам не удалось установить каких-либо зависимостей (как и профессору H.Selye), позволяющих связать тип действия препарата с ожидаемым и даже полученным эффектом.
Теперь, по прошествии многих лет, становятся понятными эти результаты, поскольку «цена» эффективности была продиктована не специфичностью действия препарата, а способностью клетки «оплатить» предъявленную ей «цену» из арсенала собственных резервов.
Тогда, в группе самых эффективных препаратов (креатинфосфат, неотон, глюкозо-6-фосфат) и смоделированных состояний (гипертрофия рабочего миокарда) максимальный прирост времени переживания тотальной ишемии сердцами крыс, на которых проводились испытания, составил лишь 10-15 минут при +35 С и 50-60 минут - при +16+18 С.
Ретроспективно можно сказать, что такова была косвенная характеристика объема резервных возможностей кардиомиоцитов и, конечно, миокарда.
Пример 9. Гипоксическая и циркуляторная гипоксия. Следующее состояние, с которым в практике часто приходится сталкиваться - это гипоксическая и циркуляторная гипоксия. Ее действие менее жестоко по отношению к системам гомеостатической регуляции, поскольку их редукция здесь весьма относительна и определена относительным же дефицитом вещества, энергии и информации. Сопровождается она теми же реакциями, что и описанные выше и по тем же схемам, завершаясь, как правило, состоянием компенсации этого дефицита гиперфункцией и умеренной гипертрофией (согласно правилу В.Ф. Фролова) за счет успешного включения механизмов долговременной адаптации (по Ф.З.Меерсону), изменения содержания циклических нуклеотидов[58], активации ДНКазы, синтеза РНК[59],[60], белка [61], либо активацией обменных процессов[62]. В большинстве же случаев гипоксическая и циркуляторная гипоксия могут даже вызвать эффект усиления противоишемической устойчивости за счет формирования «системного структурного следа» (по Ф.З. Меерсону) и/или «тренинга»[63],[64],[65], либо за счет активации обменных и регенераторных процессов[66], связанных также с активацией процессов репликации ДНК и синтеза РНК.
Пример 10. Состояние денервации, которое часто сопутствует хирургическому лечению, особенно, если речь идет об операциях, связанных с трансплантацией органов (в частности, - сердца), также травмирует систему внутриклеточной гомеостатической регуляции[67],[68], хотя и в наименьшей степени. Денервация лишает кардиомиоциты только одной составной части в системе нейрогуморальной регуляции: нервной регуляции.
Тем не менее, и денервация накладывает свой губительный отпечаток на состояние системы клеточной регуляции гомеостаза. И это находит свое отражение в неспецифической компоненте реакций отторжения, в усилении «нормальных органных выделений»[69] после операций, о чем косвенно свидетельствует усиление активности сывороточной КРК-МВ[70],[71], и активация регенераторных процессов.
Обобщая вышеизложенное, следует особо подчеркнуть значение систем регуляции внутриклеточного гомеостаза в формировании устойчивости клеток к внешним воздействиям. Именно они (системы), а точнее - их взаимоотношения, определяют и формируют функциональный и энергетический резерв клеток.
Чтобы увеличить такой резерв, и повысить таким образом устойчивость клеток к внешним неблагоприятным действиям, необходимо активизировать механизмы внутриклеточной регуляции гомеостаза с обязательным привлечением систем более высокого уровня: тканей, органов, систем органов. А это возможно только при условии детального понимания характера и глубины «возмущения» этих систем в ответ на какое-либо действие (их редукцию или иное вмешательство).
Пример 11. Исследуя резервные возможности миокарда сердец собак и крыс[72] к каким-либо воздействиям (ишемия, фармакологическая кардиоплегия, предварительная и срочная фармакологическая подготовка, холод, изоляция, облучение и т.д.), нам удалось установить, что энергоёмкость, а следовательно, объём термохимических процессов, составляющих естественную реакцию клеток на практически любое совершаемое и допустимое экспериментом сколько-нибудь разумное действие, практически одинаково. Она составляет в норме 4-4,5 Дж·г-1 с момента начала действия и до момента завершения физиологического ответа и совершается со скоростью 0,9-1,2 Дж·мин-1·г-1. При этом, было весьма убедительно доказано, что основным механизмом привлечения резервов и резервной устойчивости к таким действиям и стрессу вообще является снижение так называемого «порога критического содержания АТР» в миокарде с 2-3 мкМ·г-1 до 0,5-0,8 мкМ·г-1.
Поскольку любое вмешательство в естественный гомеостатический баланс в обмене веществом, энергией и информацией, а уже тем более такое грубое вмешательство, как уничтожение одного или нескольких уровней регуляции (что мы могли наблюдать в приведенных выше примерах), не может не вызвать острого компенсаторного перераспределения соотношений оставшихся уровней управления. Возникает вполне естественный неспецифический ответ реакцией «возмущения» неравновесной системы. Любое же «возмущение», как мы могли убедиться, имеет свою конкретную «цену» (правило Н.А.Онищенко), выраженную в дефиците энергии и дополнительном её расходовании на восстановление нормального баланса «нормы реакции». А потому эффективность вмешательства прямо зависит от объема резервных возможностей[73].
Пример 12. Говоря о кардиомиоцитах и миокарде, можно с уверенностью добавить, что их резерв (в том числе и к ишемии) есть не что иное, как способность сохранения гомеостаза в экстремальных условиях гипер и гипофункционирования через перераспределение метаболических энерготрат и потери энергии во внешней функции, диссипации и т.д.63
Срочная и долговременная фенотипическая адаптация
в системе регуляции клеточного гомеостаза
Взаимодействие всех названных выше систем регуляции гомеостаза на уровне клетки, в конечном смысле, формирует устойчивость организма, систем органов, собственно органов, тканей и самих клеток к внешним воздействиям, являясь одним из наиболее важных и интересных предметов для изучения. Ведь именно на уровне клеток решается вопрос об адекватности этого формирования, координации и исполнения норм гомеостатических реакций всего организма в целом. Не что иное, как клетка, наделённая эволюцией полным набором инструментального обеспечения норм реакций, составляет систему тканей и органов, обеспечивая гомеостатическое регулирование на уровне всего организма всеми известными путями, рассмотренными выше:
1. изменением концентрации субстратов;
2. изменением концентрации эффекторов (активаторов и ингибиторов);
3. индукцией или репрессией физико-химических преобразований;
4. индукцией или репрессией деятельности мултиферментных систем;
5. центральной регуляции высшей нервной деятельности;
6. нейрогуморальной и, безусловно,
7. межуточными отношениями.
Повышенный интерес к регуляторным системам вполне закономерен ещё и потому, что, связан с возможностью идентифицировать главные и по своей природе неспецифические (типические) реакции каждой клетки фактически на любое воздействие, которое всегда завершается специфическим (характерным для каждой клетки) физиологическим ответом, составляющим суть её обратной связи со своими «подчинёнными» органоидами и органеллами, с вышестоящим ей иерархом (организмом) и эквипотенциальными ей же участниками общего процесса (другими клетками), будь то гормон синтезирующие клетки, сократительные структуры или нейросистема.
В любом случае, описывая адаптивные процессы, мы подразумеваем взаимодействие неравновесных систем, какими являются организм и клетка, с внешней для каждой из них средой. Стоит только выделить способы акцепта и модуляции получаемой ими информации (как мы это пытались сделать при описании органоидов), становится достаточно просто проследить последующие трансформации уже известных нам способов преобразования любой поступающей информации в дееспособную, которая может нарушить и нарушает установившееся и поддерживаемое гомеостатическое неравновесие.
Такое неравновесие в физиологии и биофизике принято называть «неравновесием покоя»[74], собственно и составляющим на этом уровне организации живой материи суть нормы реакций. А потому, теперь её можно уверенно назвать «нормой метаболической (гомеостатической) реакции» или «нормой метаболической (гомеостатической) регуляции».
Однако если на уровне органоидов и органелл, нам удалось объяснить реакцию «возмущения» со стороны элементарных участников, опираясь на «теорию возмущения молекулярных орбиталей», вызывающих смещение содержания или активностей реагентов в исполнении норм реакций, то теперь, перейдя на более высокий уровень в организации «живой материи» - клетке, мы сталкиваемся с новым типом нормы реакции, где принимают участие и органоиды, и органеллы, располагающие собственным инструментальным обеспечением и собственными исполнительными инструментами, находящимися в цитоплазме. При этом цитоплазматические процессы исполняются фактически такими же инструментами регулирования, что и в органоидах, но уже выступают с ними как равноправные (эквипотенциальные) партнёры.
Это усложнение не так тяжело для анализа, как представляется, если вспомнить его побудительный мотив. А им всегда является, как мы установили, термодинамическая целесообразность (эффективность функционирования). Это значит, что только увеличение энтропии неравновесной системы способно активизировать процессы поддержания её энтальпии (если речь идет о гомеостазе). Тогда в любом действии, каким бы оно ни было и какие бы инструменты оно ни привлекало, выделяются три и только три составляющие, из которых слагается вся его сложность и вся его специфичность:
-
действие
-
акцепт
-
ответная реакция
Тогда и само действие имеет только три возможности:
-
на обмен веществом,
-
на обмен энергией
-
на обмен информацией
Тогда ишемия, гипоксия, голодание, холод, гипер- и гипотермия, токсикоз, травма, действие фармакологических препаратов и т.д. и т.п. лишь изменяют качественно и количественно характер такого обмена, смещая соотношение участников, исполняющих ту или иную норму реакции, кем или чем бы они ни были.
Возвращаясь к схематическому представлению функциональных отношений каждого из «самостоятельных» участников единой биотехнической системы «клетка-организм» (рисунок 23), абстрагируем этих участников до условного отношения:
1. гомеостатическая система (например, клетка), обладающая некой энтропией [S1] и
2. внешняя ей среда (в этом случае - организм), имеющая свою энтропию [S2]
Ввиду полуоткрытости гомеостатической системы клетки S1 и её устойчивым неравновесием по отношению к организму S2, между ними, как партнёрами, происходит постоянный обмен неким веществом m и энергией E, составляющих суть обменной информации I в балансе установленной нормы их реакции, представленной постоянной k (рисунок 24 А):
[I][S1][I] -1 [S2]-1= k = S1·S2-1
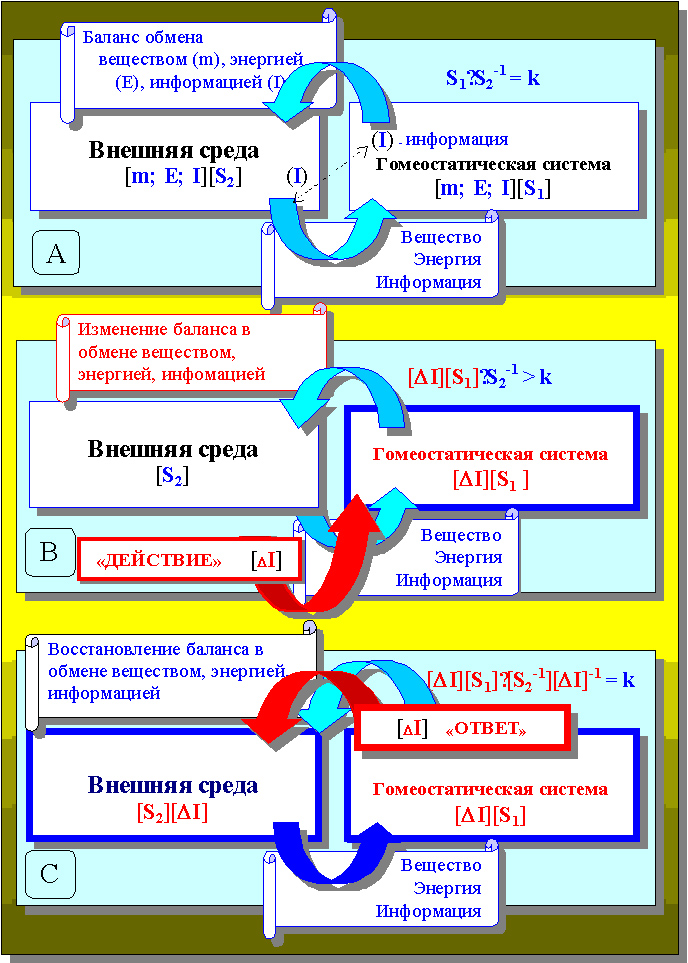
Рисунок 24.
Реакция «возмущения» условной гомеостатической системы,
находившейся в состоянии покоя (А)
в ответ на изменение условий «внешней среды» (В)
и формирование физиологического ответа (С)
Тогда любое совершённое действие извне со стороны внешней среды или просто изменение состояния одного из двух этих участников непременно вызовет смещение одного из участников отношения S1·S2-1.
Назовём это совершенное действие или изменение состояния величиной изменения обменной информации DI (рисунок 24 В), если оно было совершено извне и по отношению к клетке:
[DI][S1]·S2-1 > k
Необходимость исполнения отношения k нормы реакции S1·S2-1 наших партнёров будет безусловно сопровождаться изменением состояния S2 вплоть до момента восстановления нормального отношения k через изменение обменной информации на величину количественно равную величине DI, которая и составит адекватный действию физиологический ответ системы S1 системе S2, как своему «верховному иерарху» (рисунок 24 С):
[DI][S1]·[S2]-1 [DI]-1 = k
Результирующий эффект от представленных сейчас событий очевиден, - это восстановление отношений S1·S2-1 в исполнении нормы их реакции:
S1·S2-1 = k
Если действие прекратиться, процесс преобразований повторит только что выполненные действия, а это означает, что описание процесса адаптации клетки, обозначенное как «действие» может подразумевать любое действие, определяемое как некий «стресс-фактор», вызывающий смещение отношений или активностей участников исполнения норм метаболической регуляции. Обозначение же «ответ» сразу приобретает форму любых клеточных реакций, последовавших за этим. Набор таких реакций в клетке - есть суть реакций фенотипической адаптации, выражающиеся в физиологическом ответе на какое либо действие[75].
Согласно «Основным положениям концепции В.В. Парина и теории Ф.З. Меерсона», процесс фенотипической адаптации складывается из четырех основных звеньев или этапов (рисунок 25):
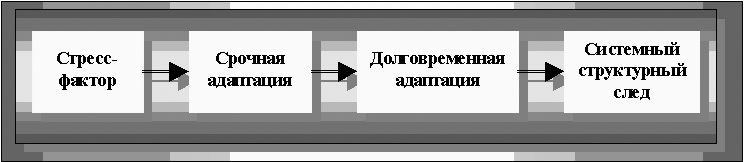
Рисунок 25. Последовательность стадий фенотипической адаптации по Ф.З. Меерсону
1. стресс фактора, которым в приведённых выше примерах выступает дефицит кислорода, дефицит субстрата, избыточная нагрузка, токсикоз, действие различных фармакологических препаратов и т.д.;
2. этапа срочной адаптации;
3. этапа долговременной адаптации, завершающихся
4. формированием «системного структурного следа».
Возвращаясь к схеме фенотипической адаптации в теории профессора Ф.З. Меерсона (часть 1, глава 3), можно выделить главное связующее звено в цепи рассматриваемых событий. Это – «нарушение гомеостаза или его предвестники» (рисунок 26). Выделение этого участка представляется очень важным, т.к. позволяет чётко определить центральное звено адаптивного процесса и установить его неспецифические черты.
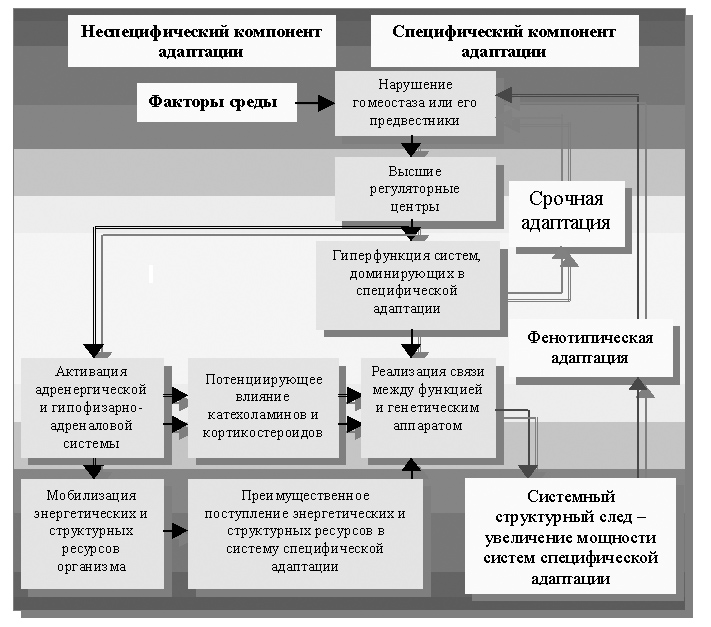
Рисунок 26. Механизм фенотипической адаптации по Ф.З. Меерсону
Та часть процессов, которая находится в левой части рисунка 26, была названа профессором Ф.З. Меерсоном «неспецифическим компонентом» фенотипической адаптации. Правая же часть рисунка 26 получила название «специфического компонента». К «неспецифическому компоненту» автором были отнесены только процессы энергообразования, синтеза белка, симпатоадреналовая и гипофизарная регуляция.
Трудно согласиться вполне с таким упрощённым разделением специфической и неспецифической компонент реакции адаптации, поскольку оно не включает в себя очень важные звенья всей цепи неспецифической регуляции. Это: нарушение гомеостаза (реакция «возмущения»), гиперфункция (физиологический ответ). Их тесная взаимосвязь определена представлениями классической патологической физиологии и представлены принципом В.А. Фролова «о гиперфункции»[76].
Скорее всего, следовало бы в приведенной схеме в качестве неспецифической компоненты реакции выделить только «стресс-фактор», хотя и его «неспецифичность» более чем условна. Однако такие важные заключения не должны оставаться априорными и требуют своего обязательного комментария.
Дело в том что, выделив, или точнее, разделив специфическую компоненту реакции на внешнее воздействие - «стресс-фактор» и «неспецифические составляющие», а также установив реальный орган исполнения регуляторной функции – мембрану клетки, не хочется ограничиваться только описательным характером адаптивных процессов. Как правило, именно это вносит серьёзную погрешность в понимание механизмов реализации защитных свойств всех систем регуляции (поправка Е.И. Чазова), а значит, не позволяет правильно разработать пути их усиления или практического использования.
Только подойдя к субклеточному (или мембранному) уровню в трактовке адаптивных процессов, становятся очевидными реальные участники, или «материальные» части процесса, собственно определяющие вторую и главную неспецифическую компоненту адаптивно-приспособительных реакций.
Это структуры, входящие в состав цитоплазматических мембран и некоторые их свойства:
1. трансмембранные ферментные структуры: рецепторные зоны аденилатциклазы, гексокиназы, иммуноферментов или R белков и т.д.;
2. трансмембранные коммутативные структуры ионоселективных каналов К+-Na+АТФ-азы, системы Na+-Са++-обмена и элементы диффузионных транспортных систем (механизма облегченной диффузии);
3. производные трансмембранного потенциала (φ): электрическое, магнитное, электромагнитное и электростатическое поле мембраны.
Во всех клеточных реакциях эти участники принимают самое непосредственное действие, а значит и сам «ответ» со стороны клетки должен включать в себя этих участников, кем бы или чем они ни были. Необходимо только попытаться уточнить центральных посредников или мессенджеров[77], способных преобразовать получаемую ими информацию до уровня её дееспособности.
Для лучшего понимания идей сугубо теоретического свойства, но способных быть применимыми для решения прикладных задач, постараемся представить себе всю последовательность событий, возникающих в некой обобщённой клетке при каком-либо действии. Для этого обратимся непосредственно к сути механизмов акцепта совершаемого действия.
Акцепторный механизм исполнения реакций срочной адаптации
Чтобы не нарушать избранный нами путь изложения, не будем уклоняться от использования в качестве иллюстрирующего объекта сердце и кардиомиоцит, а также те конкретные примеры из опыта наших коллег и собственного опыта, которыми мы уже имели возможность воспользоваться. Тем более что именно этот опыт и эти обобщения привели к пониманию механизмов гомеостатической регуляции и возможностей осознанного применения этих знаний на практике[78].
Если безоговорочно принять схему профессора Ф.З. Меерсона за основу, то внутриклеточная регуляция, построенная только по принципу неспецифического ответа, выпадает из неё. В связи с этим, чтобы не заниматься излишней модификацией схем фенотипической адаптации, что не является задачей этой работы, возьмем за основу другую, несколько более простую схему (рисунок 27), что видится вполне правомерным для перечисленных выше примеров[79].
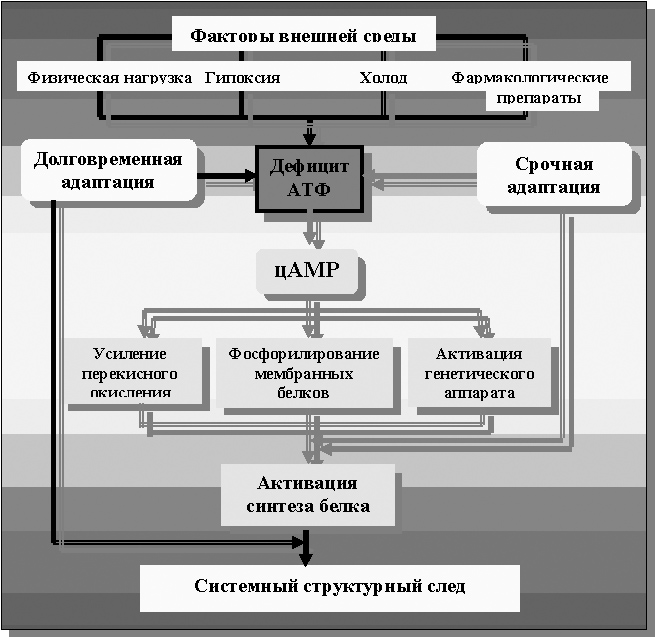
Рисунок 27. Схема клеточного звена фенотипической адаптации по В.И. Кирпатовскому
Эта схема была принята профессором Ф.З. Меерсоном[80] ещё в ранних его работах, от которых сегодня профессор отдалился. Она была чуть более простой, чем приведённая выше, но, по сути, более стройной, чем схемы, приведённые ранее. Претерпев дополнения, внесённые профессором В.И. Кирпатовским ещё в его кандидатской работе[81], означенная схема достаточно чётко указывает на искомое существо неспецифической компоненты адаптивного процесса на клеточном уровне – это «дефицит энергии» (правило Н.А. Онищенко).
В связи с этим, дальнейшее изложение в этой главе будет построено по принципу рассмотрения «изолированной»[82] регуляции клеточного гомеостаза. Это позволит более точно рассмотреть неспецифическую компоненту (или компоненты) в сложной цепи биохимических процессов, происходящих в клетках.
К первому и наиболее важному звену в цепи последующих рассуждений следует, на наш взгляд, отнести факт внесения в схему такой составляющей как дефицит энергии (или дефицит АТР).
Между тем, известно, что состояний, связанных с хоть сколько-нибудь продолжительным во времени реальным дефицитом АТР (если это соизмеримо с жизнью), не существует и не может существовать, т.к. абсолютное содержание АТР является величиной регуляторной, представляющей сиюминутную материальную суть баланса между расходованием и синтезом АТР и определяющей, таким образом, только результирующую от реального хода событий[83].
Пример 13. Абсолютное содержание АТР может быть низким при высокой скорости его синтеза и высокой скорости расходования, например, у здорового тренированного спортсмена и может быть много выше в больном гипертрофированном миокарде при низкой скорости её синтеза и ещё большей не способности её адекватного использования, например, при одновременном действии на сердце предварительными средствами противоишемической защиты: α-токоферолом и интраоперационно кардиоплегическим раствором и холодом[84], когда суть «критичности» состояния систем регуляции гомеостаза при многофакторных нагрузках связана с «аффективностью» факторов разного типа действия.
Объективно регулятором исполнения «нормы реакции» фосфорилирования АDР и дефосфорилирования АТР является скорость их синтеза и расходования, связанная с отношением: [АDР][P][АТР]-1 и [Кр][P][КрР]-1 и условиями исполнения этих отношений[85],[86],[87]. Тогда увеличение абсолютного содержания АТР только регламентирует скорость его фосфорилирования, а уменьшение[88], - увеличивает.
Вместе с тем, над этим первым реальным «исполнителем» адаптивных процессов и в полной зависимости от него стоят такие регулирующие факторы (условия исполнения) как:
1. уровень поляризации мембран, который нам очень хочется назвать «физическим мессенджером» или фактором прямого физического регулирования, определяемым величиной трансмембранного ионного градиента и, соответственно, интенсивностью АТР зависимых калий-натриевого[89],[90],[91] и кальций-натриевого обмена[92],[93],[94];
2. активность (содержание) внутриклеточного кальция, получившего имя «ионного мессенджера» и зависящая от интенсивности работы АТP- и Ca++-зависимых АТPаз[95],75,76;
3. аденилатциклазы[96],[97], мембраносвязанной структуры действительного «биологического мессенджера», регулирующей опосредованно:
· скорость исполнения норм реакции [АDР][P][АТР]-1 и [Кр][P][КрР]-1;
· скорость трансмембранного ионного транспорта;
· состояние кальций-связывающих и кальций-транслоцирующих систем.
Значит, изменение состояния любого из перечисленных компонентов может вызвать изменение содержания АТP, как регуляторной субстанции, т.е. создать её условный дефицит («дефицит АТP» по Н.А. Онищенко). Равно, и наоборот: возникший по каким-то причинам дефицит АТP изменит состояние названных участников, «запустив» циклически замкнутый процесс последовательных возмущений.
В связи с этим, очень важным на начальном этапе звеном реакций регуляции клеточного гомеостаза нам бы хотелось изобразить (схематически) некий круг этих взаимоотношений (рисунок 28), пока только на уровне мессенджеров, которые в последующем встанут во главу общей блок-схемы всей биотехнической системы контроля и регуляции клеточного гомеостаза.
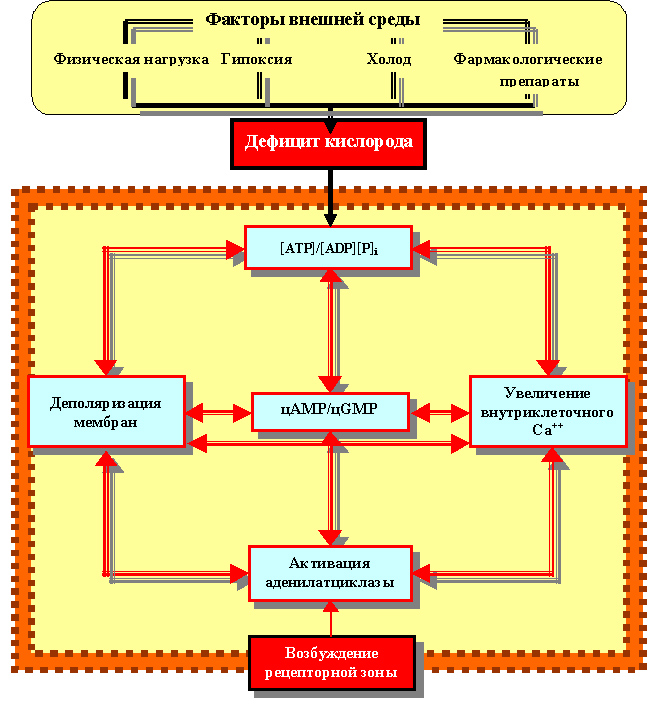
Рисунок 28. Акцепторные отношения мессенджеров в системе регуляции гомеостаза клетки (блок-схема)
Такое взаимоотношение определено строгим набором типических (неспецифических) биохимических реакций, возникающих (или активизирующихся) в ответ на изменение условий баланса в обмене веществом, энергией и информацией. Именно поэтому оно было описано таким понятием как «акцепторный механизм» клетки[98] для совершения такого обмена (вещества, энергии и информации). Физически названный тип акцепторного механизма «привязан» к клеточной мембране и способен реагировать на информацию, воспринимать её, модулировать до уровня дееспособной, превращать в набор биохимических процессов, создав возможность и условия для адекватного ответа.
Механизмы акцепта информации
В качестве «возмущающего» гомеостаз клетки «стресс-фактора» мы включили в схему наиболее вероятные для медицинской практики ситуации:
1. дефицит кислорода, характерный для многих нозологий, патогенез которых вызван, сопровождается или связан с состоянием ишемии или гипоксии;
2. деполяризацию мембран (искусственную или возникшую в общем генезе заболевая), являющуюся весьма частым (неспецифическим) следствием введения ряда фармакологических и химических агентов[99];
3. возбуждение рецепторных зон ферментов, их комплексов и систем, характерное для использования широкого спектра фармакологических препаратов и интермедиатов.
Теперь необходимо более подробно проанализировать цепь изображенных на рисунке 28 взаимоотношений участников акцепторного механизма в сложной системе регуляции клеточного гомеостаза.
Если рассматривать изолированную клетку с избранной нами позиций физики неравновесных систем и «Идеи единого гомеостатического устройства», то она представляет собой такую неравновесную систему, которая, обмениваясь с внешней для неё средой веществом, энергией и информацией, обеспечивает самостоятельное поддержание этого неравновесия60,75, т.е. в полной мере отвечает всем тем требованиям, которые предъявляются к «независимым» обладателям гомеостаза.
Тогда, воспроизведенный на рисунке 28 акцепторный механизм является для клетки атрибутом, не только служащим для передачи информации о состоянии внешней ей среды, но и для управления скоростью обмена с ней (со средой) веществом и энергией, составляющих суть обратной связи (в соответствии теории И.И. Шмальгаузена). Цепь же таких взаимоотношений хорошо известна.
Так увеличение активности аденилатциклазы приводит к ускоренному гидролизу АТР с образованием циклической формы АМР – цАМР22,24 (рисунок 29 А).
Среди цАМР-зависимых реакций необходимо выделить активацию целого каскада протеинкиназ (ПК-1, ПК-2, ПК-п, рисунок 29 В), ответственных за фосфорилирование ионных каналов79,[100], некоторых белков22,24,75, митохондрий75,[101],[102], рибосомального аппарата14,24,79, собственно плазмолеммы23, [103] и некоторых других ферментных систем79, механизм активации которых представлен на рисунке 29 С.
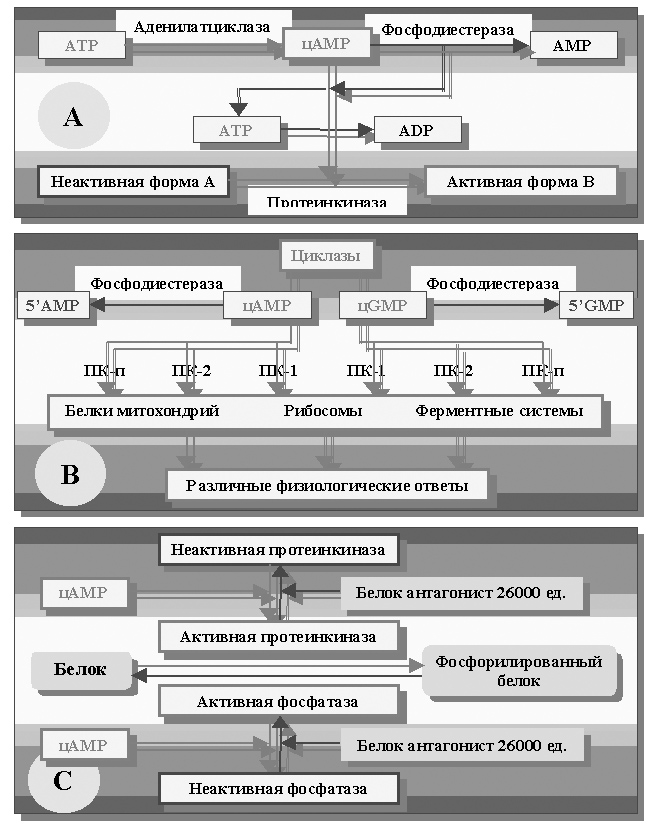
Рисунок 29. А – блок-схема регуляторной активности цАМР;
В – общая схема включения циклазной системы регуляции по Н.А. Фёдорову;
С – схема цАМР зависимого фосфорилирования и дефосфорилирования белков.
Таким образом, цАМР опосредованно, через систему протеинкиназ, связана с изменением и ионной проницаемости мембран, т.к. в ответ на фосфорилирование ионных каналов усиливает диффузные ионные токи. Это, в свою очередь, сопровождается деполяризацией мембран20,23,38, а значит и активацией расхода АТР (дефицитом АТР), необходимого на восстановление ионных градиентов и реабсорбцию кальция.
Здесь же важно отметить, что хотя молекула цАМР исключительно стабильна, в присутствии другого фермента - фосфодиестеразы, она очень быстро гидролизуется до АМР и поэтому живет очень не долго 22,24,79, что является важной и отличительной чертой практически любого участника оперативной регуляции.
При этом разрушение цАМР прекращает процесс фосфорилирования ионных каналов и, следовательно, устраняет пассивную диффузию ионов через ионные каналы, останавливая таким образом процесс деполяризации20,23,38. При этом формируется некий «двойной контроль» за управлением, принятый общей стратегией биологической регуляции[104] адаптивных процессов, усиливающих их эффективность.
Теперь необходимо выделить роль ионов внутриклеточного или «свободного» кальция (Са++) в описанном выше акцепторном механизме системы регуляции клеточного гомеостаза.
По мнению ряда авторов, активный внутриклеточный Са++ необходимо включить в ряд трех основных мессенджеров метаболической регуляции. Это: гормоны, циклические нуклеотиды и Са++ [105],[106]. При этом, ионы Са++ являются эволюционно даже более древним мессенджером, чем система циклических нуклеотидов, а потому занимают одно из главенствующих положений в системе регуляции клеточного гомеостаза.
Следующая связь в «акцепторном механизме» - это взаимоотношения Са++ и циклических нуклеотидов. Она довольно сложна, но весьма наглядно продемонстрирована схемой профессора Н.А. Федорова (рисунок 30), согласно которой можно проследить четкую взаимосвязь концентрации и/или активности внутриклеточного Са++ с:
1. активностью гуанилатциклазы и фосфодиестеразы;
2. соотношением активностей аденилатциклаза/гунилатциклаза;
3. с уровнем поляризации мембран через опосредованное фосфодиестеразой отношение цАМР/цGМР;
4. очевидна и прямая связь с дефицитом АТP22,24 через Са++АТР-азу и АТР - зависимый Na+-Са++-обмен;
5. с деполяризацией мембран такая связь еще более непосредственна, чем через фосфодиестеразу, т.к. при уровне деполяризации мембран -35-40 мВ открываются трансмембранные ионоселективные каналы и внеклеточный Са++ медленным током начинает поступать внутрь клетки (в цитоплазму кардиомиоцита)[107],[108],[109];
6. при уменьшении трансмембранного градиента по Na+, являющегося партнёром Na+-Сa++-обмена, происходит изменение направленности тока Са++ через эту же систему внутрь клетки88,[110].
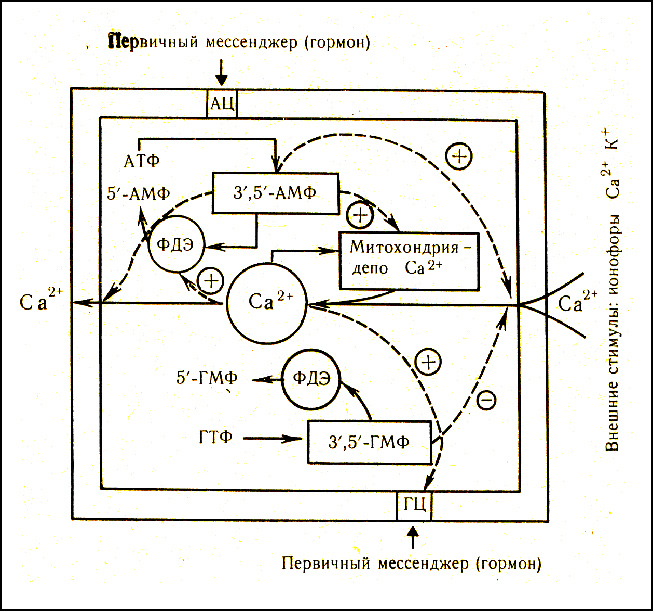
Рисунок 30. Взаимные влияния циклических нуклеотидов и Са++ на их уровень в клетке:
+ стимуляция; - ингибирование; - - - взаимные влияния
Обратные взаимодействия, т.е. Са++ – деполяризация, также понятны:
1. появление свободных ионов Са++ в цитоплазме сопровождается активацией Са++-АТP-аз и Na+-Сa++ ионообменного механизма, а следовательно,
2. перерасходом АТP на реабсорбцию Са++ обратно в «депо» и выведение его из клетки в межуточный объём 88,[111].
3. Появление же ионов Са++ в цитоплазме всегда вызывает снижение содержания АТР (дефицит АТР) и, следовательно, уменьшает поляризацию внешней мембраны клеток.
Остается проследить правомерность последней связи «деполяризация - активность аденилатциклазы». Здесь имеют место два механизма:
1. Это смещение условий равновесия исполнителей «нормы реакции» аденилатциклаза/гуанилатциклаза и цАМP/цGМP[112].
Пример 14. Что касается кардиомиоцитов, то деполяризация, вызванная, в частности, введением кардиоплегических (деполяризующих) растворов, либо электрическим током (фактором внешнего воздействия), охватывает не только изолированную клетку, т.к. она изолирована лишь условно (как мы говорили: регуляторно). В зону деполяризации попадают и хранилища эндогенных катехоламинов миокардиальной ткани, которые в ответ на такую деполяризацию должны освобождать и освобождают из своих специализированных «депо» адреналин и норадреналин ([113]), немедленно «атакующих» рецепторные зоны аденилатциклазы и смещая, таким образом, условие равновесия исполнителей «нормы реакции» аденилатциклаза/гуанилатциклаза и цАМP/цGМP[114].
2. Вторая взаимосвязь носит в большей степени гипотетический характер, хотя теоретически она более убедительна, чем первая. Хорошо известно, что деполяризация мембран вызывает увеличение их толщины[115]. Это происходит за счет изменения электростатических сил «сжимающих» липидные «обкладки» мембраны. Отрицательные заряды фосфолипидов, отталкиваясь друг от друга, перестают испытывать противодействие «сжимающих» сил, и толщина мембраны пассивно увеличивается[116] за счёт сил Ван-дер-Ваальса. Это факт.
Изменение же физического состояния мембраны не может не оказывать влияния на её структурные составляющие[117], включая интегральные белки[118]. Функциональная же активность таких белков, во многом определена электростатическими и гидрофобными связями с их липидным окружением[119],[120] в составе мембран[121], составляющих суть пятеричной формы кооперации. Это гипотеза.
3. Аденилатциклаза, являясь интегральным мембранным белком, теоретически способна изменить свою активность за счет конформации тесно связанного с ней гидрофобными и электростатическими силами[122] липидного окружения[123], хотя прямых указаний на это в литературе найти так и не удалось.
4. Из числа описанных выше взаимодействий, в акцепторном механизме адаптивных реакций взаимоотношение «аденилатциклаза – деполяризация» менее всего изучено, но легко объяснимо нормой этих отношений:
[аденилатциклаза]/[фосфодиестераза]↔[протеинкиназа]/[фосфатаза] ↔ степень фосфорилирования трансмембранных канальных белков ↔ активизация и/или угнетение трансмембранных ионных токов ↔ поляризация и/или деполяризация мембран
Таким образом, схема акцепторного механизма в цепи реакций регуляции клеточного гомеостаза, изображенная на рисунке 28, может быть признана вполне корректной. Уточнений же заслуживает лишь указанная выше связь «деполяризация ↔ аденилатциклаза».
Исходя из факторов, включенных в «акцепторный механизм», можно попытаться проследить дальнейшую цепь событий от момента действия «стресс-фактора» ↔ «акцепта информации» ↔ «модуляции информации» ↔ «увеличение энтропии системы» ↔ «реакция клетки» («реакция возмущения» гомеостатической системы клетки), до момента завершения этой реакции, где возможны два варианта[124]:
1. При максимальной редукции системы клеточной регуляции - до состояния «срочная адаптация».
2. При наличии кислорода, субстрата и времени – до состояния «долговременная адаптация».
При сохранности систем регуляции высокого уровня организации (тканевой, органной, систем органов), а это важно предусмотреть в ситуациях с предварительной фармакологической подготовкой пациентов при исходной хронической или приобретенной патологией (в частности, при изначальном ишемическом повреждении миокарда) - через срочную и долговременную адаптацию до состояния – «формирование системного структурного следа»[125].
Механизмы акцепта вещества, энергии и информации
релятивистского свойства
По существу, вещество, энергия и информация релятивистского свойства, поступающая вместе с волновыми электромагнитными и гравитационными потоками, если она является проникающей, способна реализовать те же механизмы и привлечь (возмутить) тех же участников акцепторного процесса. Разница состоит лишь в том, что сам акцепт таких потоков способен быть осуществим не на уровне субстратного, ионного и иного химического регулирования через изменение баланса «норм реакций», а с уровня возбуждения молекулярных орбиталей элементарных составляющих собственно участников исполнения нормы той или иной реакции.
В этой непривычной последовательности действия активность участников акцепторного механизма способна меняться изнутри и последовательно: с уровня элементарных отношений через субмолекулярные связи, межмолекулярные и композитные взаимодействия внутри макромолекул и сложных композитов, их комплексов и систем. Такой акцепт принципиально возможен и не требует какой-либо специальной программы действия, используя для этого уже подготовленный материальный инструмент, исторически и эволюционно способный принять информацию, модулировать её до состояния дееспособности, т.е. достичь реального физиологического ответа.
Суть такой модуляции сводится лишь к преобразованию высоких частот условного релятивистского потока малых масс в низкие частоты автоколебаний акцепторов большей массы, способствующих адаптации сверхскоростей к скоростям нормальных химических и электрофизиологических процессов, как это происходит с хлорофиллом при фотосинтезе или родопсином при светорецепции.
Тогда, по сути, эффекты срочной и долговременной адаптации могут быть достигнуты точно также как и описанные выше, т.е. через использование баланса реагентов при исполнения «норм реакций», контролируемых теми же участниками акцепторного механизма.
При этом очень важен тот факт, что чувствительность такого акцепта и механизм этой чувствительности демонстрируют уникальное своё совершенство на непосредственном стыке живой и неживой природы в едином гомеостатическом мире.
Физиологический ответ как завершающая фаза реакций адаптации
Не будет лишним еще раз подчеркнуть, что описанный выше «акцепторный механизм» и способы акцепта информации являются главным атрибутом в системе регуляции гомеостаза на уровне клеток. Лучше сказать: «ключевым механизмом акцепта информации на уровне клетки в общем для неё балансе обмена веществом, энергией и информацией с внешней средой»[126], механизмом её (информации) преобразования (модуляции) до уровня дееспособной. Т.е. способной быть «воспринятой» клеткой, а значит, способной вызвать «физиологический ответ»[127], выраженный в адекватном изменившемуся балансу в обмене веществом, энергией и информацией, изменении специфической функции клетки.
Из этого же следует, что не всякая информация для клетки может оказаться «дееспособной». Т.е. не всякая информация может служить для неё информацией.
Отсюда можно сделать два важных, как нам представляется, вывода:
1. Информация, не способная вызвать «физиологический ответ» со стороны клетки, не может быть названа для неё информацией;
2. Любое действие, которое способно вызывать «реакцию возмущения» со стороны клетки, может быть названо для неё информацией и всегда будет сопровождаться изменением баланса в обмене веществом и энергией, последовательным увеличением энтропии, дефицитом АТР, компенсируемым изменением скорости дополнительного производства энтропии и физиологическим ответом «обратной связи» на данную информацию с единственной целью восстановления рационального неравновесия.
Теперь рассмотрим такой феномен для клетки (кардиомиоцита), находящейся в условиях сохранной нейрогуморальной, системной, органной и тканевой регуляции. Такой феномен можно легко иллюстрировать частным примером:
Пример 15. Если в качестве раздражителя миокарда (возбуждающей информации), нарушающего установившееся равновесие в обмене веществом и энергией, использовать одиночный электрический импульсный сигнал, то в ответ на такой сигнал (электрическая деполяризация) произойдет акт: сокращение - расслабление, характеризующий не только активацию или «возмущение» участников акцепторного механизма клеточной регуляции, но и его (ответа) завершение - расслабление и реполяризация мембран (рисунок 31).
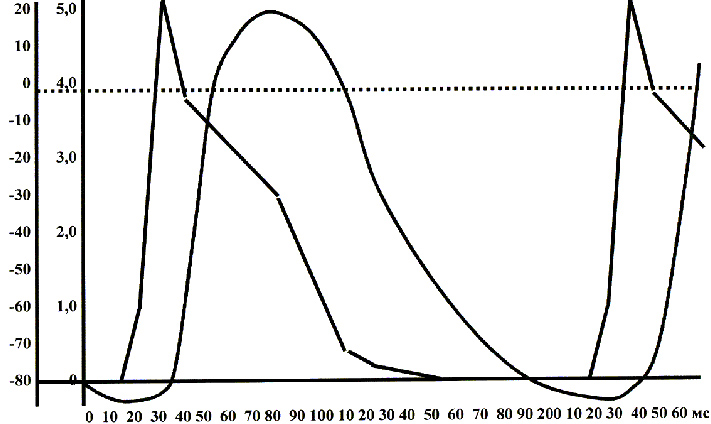
Рисунок 31. Потенциал действия и механограмма (тоническое напряжение)
рабочего миокарда сердца крысы (экспериментальные данные):
мембранный потенциал в мВ;
механограмма в г силы / г массы;
время в мс
Прокомментируем: электрический импульс, распространяясь по внешней поверхности клеточных мембран (межклеточные пространства) вызывает частичную деполяризацию плазмолеммы (в данном конкретном случае - надпороговую). При уровне поляризации мембран -40-50 мВ запускается быстрый натриевый и быстрый калиевый ток[128],[129] - мембрана деполяризуется. При уровне деполяризации -35-40 мВ запускаются медленные кальциевые токи[130],90,103,[131] - в саркоплазме появляется «свободный», не связанный кальций.
В ответ на деполяризацию и кальциевый ток, происходит высвобождение кальция из терминальных цистерн саркоплазматического ретикулума: «Са-залп» (по Ф.З. Меерсону). Все это, как уже отмечалось, требует усиленного расхода АТР на реабсорбцию кальция обратно в «депо»88,103. Содержание его в цитоплазме падает, вызывая быстро компенсируемый, но все-таки дефицит АТР, т.е. начальное увеличение скорости расхода, а затем, и скорости синтеза АТР.
Несмотря на относительную скоротечность этого процесса (100-200 мс), на наш взгляд, более внимательного рассмотрения заслуживает последовательность событий и их энергообеспечение (обмен энергией).
Используя для этих целей данные Е.Б. Бабского и соавт.[132], В.А. Фролова и соавт. и R.A. Chapman et al.[133] и других исследователей, исследовавших биохимические, биофизические и физиологические характеристики миокарда в процессе сердечного цикла и объединив полученную ими информацию в общий рисунок 32, можно с большой степенью достоверности определить последовательность происходящих событий с момента деполяризации мембран кардиомиоцитов («стресс-фактор») до момента полного расслабления миокарда.
Из представленного рисунка видно, что деполяризация мембран и увеличение тонического напряжения миокарда (механограмма) сопровождаются падением АТP и КрР, увеличением площади митохондрий, усилением теплопродукции (DG) и, сначала падением, а затем существенным ростом АМР.
Это свидетельствует за то, что выход кальция в цитоплазму вызывает временное перераспределение энерготрат: ингибирование митохондриального синтеза АТР, активацию гликолиза и гликогенолиза[134].
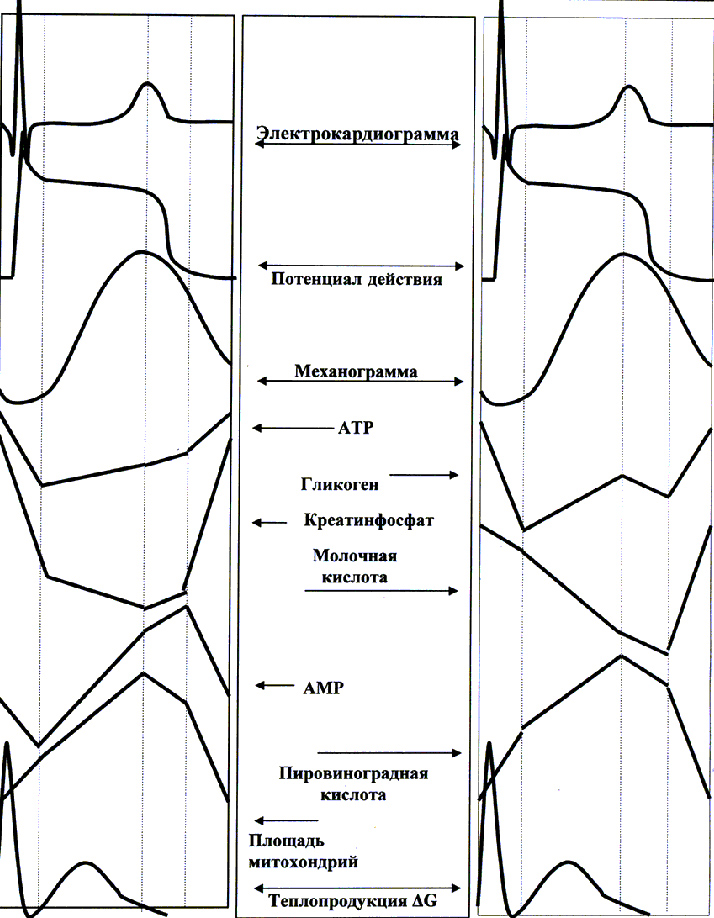
Рисунок 32. Некоторые показатели миокарда в процессе сердечного цикла (по Е.Б. Бабскому и соавт. 1975 г; В.А. Фролову и соавт. 1981-2002 г и собственным экспериментальным данным 1979-1988 г)
В дальнейшем, происходит активация митохондриального синтеза АТР: с момента максимального сокращения до полного расслабления наблюдается уменьшение площади митохондрий, увеличение уровня КрР и АТР на фоне роста запасов гликогена и нормализации содержания ПВК, молочной кислоты и активности ЛДГ.
Эти события сопровождают завершение реакции на электрическую деполяризацию. Миокард расслабляется.
Таким образом, моделью завершенной реакции адаптации, построенной по вышеописанному принципу может служить модель: «сокращение ® расслабление» кардиомиоцита:
1. фактор возмущения (информация или «стресс-фактор»),
2. активация акцепторных механизмов (акцепт информации),
3. модуляция информации (деспециализация информации[135]),
4. нарушение клеточного гомеостаза (увеличение энтропии, изменение состава внутренней среды клетки),
5. физиологический ответ (компонент обратной связи, сокращение),
6. адаптация (восстановление клеточного гомеостаза),
7. расслабление
Пример 16. Такое заключение тем более правомерно, что специальные исследования, проведенные в Екатеринбурге профессором В.С. Мархасиным[136] и профессором В.Я. Изаковым[137] показали, - что в патологически измененном миокарде, обладающим разной степенью повреждения (модификации) системы клеточной регуляции, по-разному выражена реакция изометрического сокращения. Т.е. по динамике механического изометрического сокращения (тонического напряжения миокарда) кардиомиоцитов можно косвенно судить о состоянии акцепторного механизма системы клеточной регуляции и собственно метаболических процессах, на которых он построен. Причем характеристиками чувствительности акцепторных реакций могут служить показатели процесса сокращения (время, амплитуда, полупериод), а характеристиками энергообеспечения регуляторных процессов - показатели процесса расслабления88.
Это чрезвычайно важно, поскольку последовательность событий, связанных с периодической активацией участников «акцепторных механизмов» в системе регуляции клеточного гомеостаза кардиомиоцитов и особенно, её хронотропные[138] характеристики можно с успехом проследить на примере исследования сократительной функции фрагментов миокарда в различных режимах электростимуляции или даже на целом сердце, используя не только инвазивные, но и неинвазивные методы[139].
Пример 17. Пользуясь ещё одним частным примером, рассмотрим хроноинотропные реакции миокарда, раздражаемого в режимах ступенчато нарастающей частоты электростимуляции, носящей название «лестницы Боудича». И в режиме ступенчатого убывания частоты электростимуляции – «лестницы Вудвортса»[140] .
Установившаяся амплитуда сократимости полоски миокарда при изначальной низкой частоте стимуляции свидетельствует об адаптации метаболических (регуляторных) процессов и, в частности, процессов энергообеспечения кардиомиоцитов данного устойчивого состояния (состояние срочной адаптации).
Резкое увеличение частоты электростимуляции, эквивалентное состоянию экстрасистолии, вызывает увеличение доли искусственно вызванной частотной деполяризации мембран кардиомиоцитов в единицу времени в 2 раза (стресс). В этот переходный период изменяется, соответственно, «привычная» стационарная нагрузка (удельная деполяризация - частота деполяризаций в единицу времени114 на «акцепторные механизмы» клетки, смещая их из положения равновесия (баланса) в сторону деполяризации мембран и дефицита энергии. Как результат: амплитуда сокращений немедленно падает, характеризуя несоответствие скорости синтеза АТР, скорости образования АТР. Наступающее после этого увеличение и последующая стабилизация амплитуды сокращений, является характеристикой эффективности акцепта получаемой клеткой информации, вещества или энергии в общей системе регуляции клеточного гомеостаза и характеризует скорость включения адаптивных реакций к новому режиму электростимуляции. Равно, здесь же, мы получаем информацию о резервных возможностях кардиомиоцитов в энергообеспечении такой адаптации (эффективность процесса адаптации) на «надтканевом» уровне. Это связано с тем, что данная модель не учитывает один из важнейших факторов тканевой регуляции - функциональную гетерогенность миокарда[141]. Суммарный же энергорасход является интегральным критерием. Значит, он может быть назван «универсальным» и для оценки тканевых процессов регулирования клеточного гомеостаза.
Лестница Вудвортса - ступенчатое уменьшение частоты электростимуляции, демонстрирует обратный процесс - процесс реадаптации, имеющий также большое диагностическое значение.
Резкое (ступенчатое) уменьшение частоты стимуляции позволяет косвенно, по амплитуде постэкстрасистолической потенциации, оценить интенсивность работы энергозависимых систем кальциевой транслокации в предыдущем режиме электростимуляции. Время дестабилизации амплитуды при этом является прямой характеристикой мобильности адаптационных реакций. Переходя на более низкий частотный режим электростимуляции, мы искусственно ставим кардиомиоциты в условия избытка скорости образования АТР, «возмущая» таким образом участников «акцепторных процессов» клеток изнутри. При этом можно реально наблюдать (визуализировать) процессы уменьшения скорости образования АТР по амплитуде сокращения.
Таким образом:
1. Процесс наблюдения за реальной работой системы регуляции клеточного гомеостаза не является недоступным для физиологического, т.е. оперативного контроля и может быть успешно экстраполирован по реакциям экстра- и постэкстрасистолической потенциации, а также по реакции «возмущения»: сокращение - расслабление.
2. Если реакция «возмущения» «акцепторного механизма» сопровождается для кардиомиоцитов увеличением тонического напряжения, связанного с развивающимся дефицитом АТР (скорости фосфорилирования - дефосфорилирования АТP/АDP и Кр/КрР), то завершающая реакцию срочной адаптации, фаза должна быть фазой расслабления миокарда, связанная с ликвидацией такого дефицита.
Рассматривая реакции «акцепторных процессов» в системе регуляции клеточного гомеостаза, мы умышленно выделяем ведущие ее звенья:
1. дефицит АТР, активирующий скорость синтеза энергии;
2. увеличение влияния цАМР (отношения цАМР/цGМР), усиливающего активность мембранных белков;
3. появление свободного кальция - главного мессенджера в системе регуляции клеточного гомеостаза;
4. изменение уровня поляризации мембран клеток, вносящего резкий дисбаланс в структуру энергообразования и ионного распределения.
Пример 18. Как мы показали на примере реакций сокращение - расслабление, экстра- и постэкстрасистолической потенциации, эти звенья являются пусковыми в активации процесса расслабления как физиологического ответа, завершающего цепь последовательных реакций адаптации.
Мы акцентируем внимание коллег на этом для того, чтобы подчеркнуть высокую сопряженность процессов сокращения и расслабления, их теснейшую взаимосвязь, как главную, всегда однотипную, неспецифическую форму «ответа» кардиомиоцита на действие многообразной, неспецифической для клетки, информации, которая была предварительно отфильтрована и смодулировна на уровне организма. Затем, на уровне клетки, её «акцепторными процессами» уже не столь многообразная информация претерпевает вторичную фильтрацию и модуляцию до уровня восприимчивости и превращается в дееспособную, завершающуюся типическим физиологическим ответом (специфический ответ обратной связи).
К сожалению, механизмам сопряжения процессов сокращения и расслабления кардиомиоцитов уделяется незаслуженно мало внимания, хотя именно они являются ключевым в понимании адаптационных реакций кардиомиоцитов[142]. Отсюда очень часто неверное толкование характера распределения энерготрат в сердечном цикле: максимальный энергорасход приписывают фазе изометрического[143] или изоволюмического[144] сокращения20. Это не верно.
С точки зрения поставленных нами задач, рассмотрение именно этих взаимоотношений особенно важно, т.к. оно определяет последовательность включения регуляторного звена в системе поддержания клеточного гомеостаза, устанавливает их характер. Ведь именно в период расслабления сердца, несмотря на высокий уровень энергорасхода, связанного с удалением свободного кальция и реполяризацией плазмолеммы, осуществляется восстановление содержания макроэргических соединений: АТР и КрP, запасов гликогена, происходит регенерация ультраструктур миокарда15,27,28.
Для нашего изложения важно, что активация механизмов расслабления происходит еще до момента нарастания тонического напряжения миокарда в период «возмущения» участников «акцепта» и усиливается одновременно с ним, - с нарастанием тонического напряжения миокарда, являясь конкурентной реакцией.
В чем же именно заключены эти проявления, зафиксированные нами в приведенных примерах? Прежде всего, это:
1. Увеличение уровня цАМР в ответ на активацию мембранной аденилатциклазы опосредованно через набор специфических протеинкиназ[145],22,79,87 запускает процесс реабсорбции кальция обратно в «депо».
2. Увеличение внутриклеточного содержания кальция также активирует его реабсорбцию опосредованно через систему кальмодулин ® протеинкиназы ® фосфоламбан ® АТР-азы саркоплазматического ретикулума и сарколеммы[146],[147].
3. В это же время активизируется работа натрий-кальциевого ионообменного механизма, ориентированного на удаление кальция из клетки[148].
Именно поэтому на уровне максимального напряжения миофибриллярного аппарата клеток можно наблюдать высшую точку интенсификации метаболических процессов. По всей вероятности, момент «пика» сокращения можно условно назвать равновесным состоянием между поступлением и удалением кальция «в» и «из» клетки.
В частности, между свободным и связанным с тропонином кальцием, где константа диссоциации[149] (kдис), всегда должна оставаться постоянной (если условия внешней среды, рН, в частности, неизменны)85:
kдис = [Са++][тропонин] / [Са-тропонин]
За счет активации Са-АТPаз при образовании актин-миозиновых «мостиков», вызванных взаимодействием [Са++]i с субъединицей Y тропонина свободная диссоциировавшая фракция кальция [Са++]i начинает активно удаляться из саркоплазмы (в клеточные «депо» и межклеточное пространство). Поскольку в момент сокращения скорость накопления [Са++]i в цитозоле выше скорости его удаления, подъем напряжения покоя резко нарастает (фаза сокращения). В фазу максимального напряжения миофибриллярного аппарата скорость накопления [Са++]i быстро ослабевает, уступая место скорости его удаления. При этом происходит сдвиг равновесного состояния [Са++]i [тропонин] / [Са-тропонин] в сторону удаления [Са++]i. Это вызывает интенсификацию процесса диссоциации кальций-тропонинового комплекса до исходного, равновесного состояния22,24 в соответствии с (2.2.1).
Еще более этот процесс усиливается под влиянием кальциевых антагонистов - ионов калия, активно поступающих в саркоплазму за счет интенсивной работы (именно в это время) калий-натриевой АТPазы. Сердце расслабляется, характеризуя завершение физиологического ответа клеточных реакций, направленного на поддержание постоянства внутренней среды кардиомиоцита, т.е. его клеточного гомеостаза.
Таким образом, чтобы завершить нашу схему, начатую рисунком 28 и изображающую акцепторные реакции мессенджеров, попытаемся на основании всего вышеизложенного материала изобразить это графически (рисунок 33).
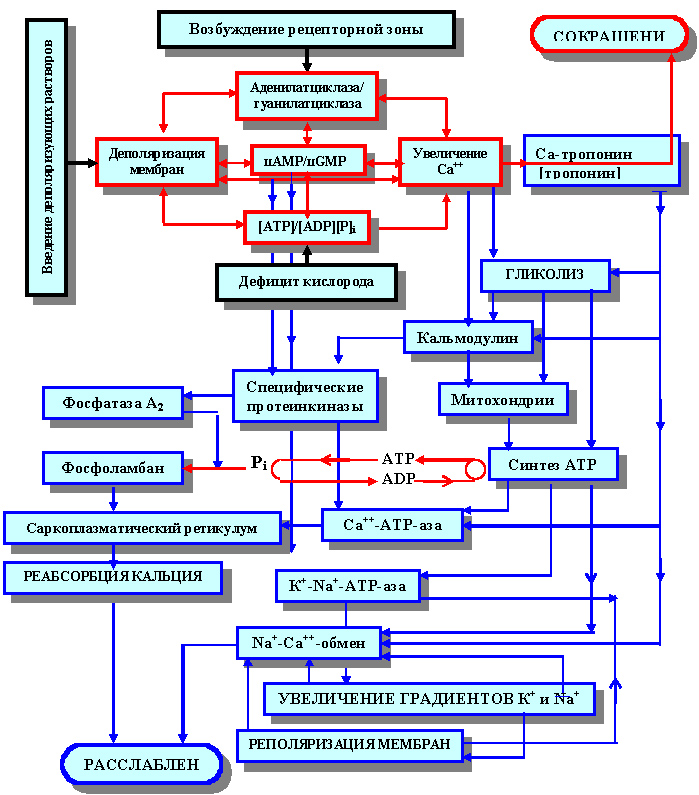
Рисунок 33. Блок-схема типической (неспецифической) реакции в системе регуляции гомеостаза клеток рабочего миокарда (кардиомиоцитов) на различные виды воздействия
В заключении нам бы хотелось еще раз выделить ключевые компоненты описанных процессов:
1. Реальный орган исполнения регуляторной функции:
- плазмолемма
2. Акцепт и модуляции информации:
- аденилатциклаза
- трансмембранный потенциал
- содержание АТP (скорость фосфорилирования / дефосфорилирования АДP/АТP и Кр/КрP)
- ионизированный кальций
- цАMP (отношение содержания цАМР/цGМP)
3. Трансляторы модулированной информации:
- ионизированный кальций
- цАМP цАМР/цGМP)
4. Регуляторы адаптивных процессов:
- кальмодулин
- фосфоламбан
- протеинкиназы
5. Донаторы энергии:
- митохондрии (протонное фосфорилирование)
- гликолиз
- свободная энергия трансмембранного потенциала (суть механизмов ионного фосфорилирования)
И хотя предложенная схема регуляторных клеточных процессов не может претендовать на обобщение механизмов долговременной адаптации[150], в общем виде последовательность происходящих событий в формировании системного структурного следа нам хотелось бы представить, немного дополнив приведенную на рисунке 25 схему, сохранив всю последовательность описываемых событий.
Схема дополнена лишь одним, но главным и как представляется, очень логически выдержанным звеном для описания процессов регулирования и «ответственного» контроля означенной последовательности, который обеспечен в клетке акцепторными отношениями мессенджеров (рисунок 34).
На наш взгляд, приведенная схема носит вполне чёткий характер и может, в определённом смысле, быть использована для системного анализа механизмов регуляции клеточного гомеостаза. Его связи с нейрогуморальными и другими процессами более высокого иерархического уровня, принимающими непосредственное участие в обмене с клеткой веществом, энергией и информацией посредством последовательного привлечения представленных механизмов. Для нас же она важна ещё и тем, что позволила на частном примере кардиомиоцита найти путь к обобщению.
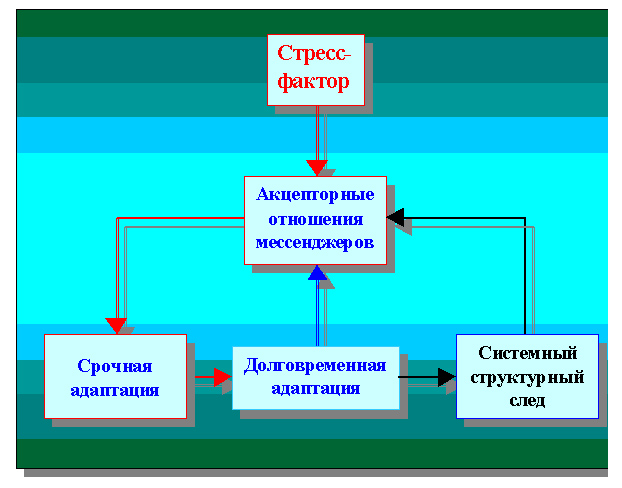
Рисунок 34. Последовательность стадий адаптивного процесса
На основании выше изложенных представлений о системах регуляции гомеостаза в свете «Идеи единого гомеостатического устройства» в общем балансе обмена веществом, энергией и информацией, особое значение приобретают такие понятия, как «резерв устойчивости» и его составляющие: «энергетический» и «функциональный» резервы.
Причем, на наш взгляд, такая категория, как «резерв устойчивости» гомеостатической системы может быть распространена только на те системы, которые неравновесны по отношению к окружающей среде с позиций термодинамики, и способны к активному поддержанию такого неравновесия. Т.е. наделены и обладают собственным гомеостазом. Т.е. - это «живые» системы. В данном контексте, смерть - есть состояние полного истощения резервов устойчивости.
Иными словами, «резервом устойчивости» любой системы можно назвать способность этой системы в полной мере использовать свой энергетический потенциал через перераспределение метаболических энерготрат и потери энергии во внешней функции, диссипации и т.д.
Под «энергетическим резервом» мы понимаем потенциальные возможности системы в энергообеспечении внешней функции, определяемой требованиями существования гомеостаза как нормы метаболических реакций.
Взаимоотношение резерва устойчивости и энергетического резерва системы определяет формирование и объем «функционального резерва».
Гомеостаз растительной клетки.
Сам факт существования растительной клетки, как «элементарного» представителя единственного продуцента в биологии трофических рядов являет собой неоспоримое свидетельство того, что эволюция живой материи в нашей планетарной биосфере носила и носит так называемый «вертикальный характер», детерминированный:
1. подвижностью
2. геофизическим местопребыванием
3. средой обитания
4. способностью не только регуляторно (рефлективно), но и продуктивно акцептовать энергию внешних (по отношению к организму) электромагнитных излучений.
В этой связи, в описании специфических особенностей гомеостаза растительной клетки в интересующем нас аспекте выявления характерных неспецифических элементов устройства, следует отметить лишь то обстоятельство, что этот гомеостаз, как свойство живого, также отвечает всем предъявляемым к нему требованиям изначального определения:
1. внутренняя среда растительной клетки, также как животной, неравновесна по отношению к окружающей её среде;
2. она также способна самостоятельно поддерживать своё неравновесие;
3. она также отделена от внешней среды очевидной границей несмешиваемых фаз (мембрана и оболочка);
4. она также обменивается с внешней для неё средой веществом, энергией и информацией, сохраняя этот баланс в течение всей своей жизни;
5. она также наделена органоидами и органеллами, исполняющими в ней отведённые роли;
6. у некоторых растительных клеток также можно определить свойство раздражимости и возбудимости;
7. некоторые из них также весьма подвижны, а большинство наделено свойством фототропизма и т.д.
Основное и определяющее отличие в том, что обладая способностью к фотосинтезу, растительная клетка способна акцептовать и преобразовывать энергию солнечного света в энергию молекулярных связей, используя для этого идентичные механизмы «возбуждения» молекулярных орбиталей.
Эта способность определяет, по своей сути специфические гомеостатические особенности, которые, при желании, можно найти в литературе, посвященной физиологии растений.
Вместо заключения
Система поддержания клеточного гомеостаза складывается из пяти основных уровней регуляции: системной, органной, тканевой и собственно внутриклеточной. Они же формируют два наиболее совершенных гомеостаза: организменный и клеточный[151], максимально защищающий от каких либо внешних трудностей гомеостаз органоидов и органелл, как главную арену исполнения нормы гомеостатических реакций, где живая материя производит свои самые сложные метаболические преобразования.
Все высшие (надклеточные) этажи гомеостатической регуляции, а равно гомеостаз организма, эволюционно были сформированы для сохранения постоянства внутриклеточной среды (клеточного гомеостаза) и взаимосвязаны с ним сложным набором причинно следственных отношений (типических реакций), включающих в себя:
1. реакцию «возмущения» при акцепте информации,
2. процессы модуляции поступающей информации,
3. характерный физиологический ответ, как элемент оперативной обратной связи для исполнения адекватной функции в исполнении общей нормы гомеостатических реакций
4. последующую адаптацию (т.е. восстановления баланса в обмене веществом, энергией и информацией).
Более того, именно взаимоотношение всех систем регуляции клеточного гомеостаза определяет и формирует функциональный и энергетический резервы клеток[152] в их адаптивности и устойчивости к внешним воздействиям, нагрузкам и т.д., т.е. к изменению условий в обмене веществом, энергией или информацией, диктуемых более высоким иерархом. Адаптивности и устойчивости к любому иному изменению «норм реакций», выраженных балансом или нормами обмена вещества, энергии и информации. Причем собственно процесс акцепта такой информации и характер реакции «возмущения» определяют ключевое звено в этом обмене, отражая, таким образом, состояние отношений всех участников исполнения условий поддержания клеточного гомеостаза[153].
Об эффективности действия акцепторных процессов и реакции «возмущения» можно судить путем непосредственного измерения её составляющих, а равно по характерным реакциям специфического (типического) ответа, как основного элемента обратной связи с вышестоящими иерархами и уровнями их регуляции, координатора на эквипотенциальном уровне и управляющего на подчинённом ему уровне. При этом эффективность функционирования всей системы может быть определена отношением физиологического ответа к суммарному расходу энергии на этот ответ.
Важно также, что информация, не способная воздействовать на систему клеточной гомеостатической регуляции, не может быть названа информацией для данной системы (для клетки).
И напротив, все то, что способно вызвать изменения в балансе обмена веществом и энергией в процессе акцепта, может быть названо информацией и всегда будет сопровождаться увеличением энтропии системы её акцептовавшей, и следовательно, завершаться физиологическим ответом[154] вплоть до восстановления начального баланса неравновесия (энтальпии) системы[155].
Пример 19. Энергетический и функциональный резерв в системе клеточной гомеостатической регуляции кардиомиоцитов и эффективность функционирования акцепторных процессов находят свое отражение в реакциях «сокращение – расслабление», реакциях хроноинотропии, поэтому их исследование (резервов) может быть проведено классическими приемами современной физиологии. В частности, по скорости сокращения миофибриллярного аппарата кардиомиоцитов можно судить о сохранности процессов сопряжения работы акцепторных реакций клетки и биохимических превращений в механический физиологический ответ. Учитывая увеличение доли искусственно вызываемой деполяризации мембран кардиомиоцитов в единицу времени в соотношении с амплитудой механического сокращения можно оценить эффективность энергетического резерва. При этом, время стабилизации амплитуды сердечного сокращения является прямой характеристикой эффективности выполненного акцепта, а также характеризует скорость включения (а значит и сохранность) реакций срочной биохимической адаптации и т.д.
Иными словами, процесс наблюдения за функционированием системы клеточной гомеостатической регуляции, её энергетическим и функциональным резервами не является недоступным для физиологического, а значит и динамического контроля и может быть успешно применен на тканевом и клеточном уровнях[156].
Главный же вывод, который хотелось бы сформулировать сейчас, ориентирован на то, что наибольший интерес для нас сегодня в оценке жизнеспособности и функциональной полноценности клеток, тканей, органов, их систем и организма в целом лежит в физиологическом или «функциональном» подходе к энергетике этих систем, трактуемых в соответствии с «Идеей единого гомеостатического устройства». Здесь, как нам кажется, можно много, интересно, плодотворно и с большой практической пользой работать.
Это тем более видится важным, поскольку подчиненность вышеописанных процессов всеобщим законам физики бытия и развития, предоставляет нам уникальный, по своей сути, шанс экстраполировать ключевые позиции механизмов неспецифического свойства, выявленных на уровне клетки, на все этажи гомеостатического устройства, отслеживая и выделяя в сравнительном анализе их особенности.
Глава 5
Гомеостаз организма
[1] Cellula, cytus
[2] В соответствии с клеточной теорией немецкого физиолога и цитолога Теодора Шванна (1810-1882) и немецкого патолога Рудольфа Вирхова (1821-1902)
[3] бактерии, простейшие, некоторые водоросли, грибы
[4] клетки, имеющие ядро
[5] безъядерные клетки
[6] митоз – (от греческого mitos - нить) основная форма клеточного деления, сущность которой заключается в равномерном (диплоидном - двойным) распределении хромосом между дочерними клетками
[7] мейоз – (от греческого meiosis - уменьшение) тип деления клеток (гаметоцитов), характеризующийся образованием из одной клетки четырёх гамет с гаплоидным (одинарным) набором хромосом
[8] или компартментация
[9] диффузия, облегченная диффузия, активный транспорт, пиноцитоз, фагоцитоз, ионообменные процессы
[10] как пример долговременной адаптации, выраженный в структурно функциональном устройстве
[11] клетки покровных тканей, эпителиев, опорные, соединительные, двигательные, раздражимые и т.д.
[12] если возможность выжить, продлив свой род и вид у одноклеточных связана с увеличением численности клеточной популяции (компенсация митотической активностью), то у многоклеточных, - с усовершенствованием защитных и поведенческих реакций.
[13] Гипотеза Андрея Сергеевича Фаминцына (1835-1918) о происхождении организмов путем их симбиоза
[14] Заварзин А.А., Харазова А.Д. Основы общей цитологии, Л.ЛГУ, 1982, 239 с., Заварзин А.А. Основы частной цитологии и сравнительной гистологии многоклеточных животных, Л.ЛГУ, 1976, Ченцов Ю.С. Общая цитология, 2 изд., М.1984; Rogers A.W. Cells and tissues. An introduction to histology and cell biology? L.1983
[15] в зависимости от уровня организации самого организма (от простейших форм до высших)
[16] межклеточное или межуточное взаимодействие
[17] это происходит уже в третьем пассаже культуры клеток (данные Института эпидемиологии и микробиологии им. Н.Ф. Гамалеи и Института вирусологии им. Д.И. Ивановского)
[18] Этот факт также можно рассматривать как аргумент в пользу единого происхождения всех клеток. С практической же точки зрения - эта «виртуальная» особенность «обратного или обращённого онтогенеза» очень активно в последние годы привлекла к себе внимание коллег (трансплантологов, гематологов, онкологов, эндокринологов и др.) при создании так называемых «высоких клеточных технологий» в заместительной терапии достаточно широкого нозологического спектра заболеваний (Материалы XIII (76) сессии общего собрания Российской Академии медицинских наук 17-20 февраля 2004 года. М, 2004).
[19] Кардиомиоцит, как объект анализа, замечателен тем, что исследование его внешней функции и метаболических особенностей, а именно, его возбудимости, раздражимости и сократительных свойств позволяет четко дифференцировать изучаемые фазы любого процесса в нормальных и патологических условиях. Тем более, что такие особенности присущи практически всем клеткам, только с разной степенью выраженности и/или развитости.
[20] Меерсон Ф.З. Теория индивидуальной адаптации к среде и профилактика стрессорных повреждений сердца. /Текст лекций// М.: УДН, 1981, 46 с.
[21] Меерсон Ф.З. Адаптация, стресс, профилактика. М. Медицина, 1981г., с.278.
[22] Саркисов А.С. (под ред.) Структурные основы адаптации и компенсации нарушенных функций./ Руководство / / М. Медицина. 1987г., 433 с.
[23] Адо А.Д. (под редакцией) Патологическая физиология. М.: Медицина, 1973, 553 с.; Фролов В.А., Дроздова Г.А., Казанская Т.А., Билибин Д.П., Демуров Е.А. Патологическая физиология. М, 2002, 707 с.
[24] Лайтфут Э. Явления переноса в живых системах. М.: Мир, 1977, 518 с.
[25] Образцов И.Ф., Ханин М.А. Оптимальные биомеханические системы. М.: Медицина, 1989, 271 с.
[26] Levy M.N., Martin P.Y. Нейрогуморальная регуляция работы сердца./В кн.: Физиология и патофизиология сердца. М.: Медицина, 1988, т.2, с.64-90
[27] Волькинштейн М.В. Биофизика, М.: Наука, 1988, 591 с.
[28] Котык А., Яначек К. Мембранный транспорт. М.: Мир, 1980, 338
[29] Ленинджер А.Л. Основы биохимии. М.Мир, 1985г., 1057 с.
[30] Keynes R.D. Ионные каналы в мембране нервной клетки./В кн.: Молекулы и клетки. М.: Мир, 1982, вып.7, с.176-190
[31] Мецлер Д. Биохимия. М. Мир, 1980г., 606 с.
[32] Cantor Ch., Schimmel P.R. (Ч.Кантор, П.Шиммел) Биофизическая химия. М.: Мир, 1985 г., т.3, 483 с.
[33] Miura D.S., Hoffman B.F., Rosen M.R. The effect of extracellular potassium in the intracellular potassium membrane potential of beating canine cardiac Purkinje fibbers. / In.: J. Gen. Physiology. 1977, 69, N 4, p.463- 474
[34] Пауков В.С., Фролов В.В. Элементы теории патологии сердца. М. Медицина. 1982г, 270 с.
[35] Фролов В.А., Пауков В.С., Казанская Т.А. и др. О некоторых общих закономерностях патологических процессов в сердце./В жрун.: Архив патологии, 1971, 1, с.14-23
[36] Фролов В.А., Дроздова Г.А., Казанская Т.А., Билибин Д.П., Демуров Е.А. Патологическая физиология. М, 2002, 707 с.
[37] Онищенко Н.А., Лубяко А.А. Противоишемическая защита миокарда кардиоплегическими растворами. / В жрун.: Вестник АМН СССР, 1985, 4, с.40-47
[38] Лубяко А.А. Противоишемическая защита миокарда кардиоплегическими растворами. Дис. ... канд.биол.наук. М. 1986г., 189 с.
[39] Скулачев В.П., Джунед Х., Брайнес А.С. Окисление и фосфорилирование в митохондриях эмбриональной мышцы. / В жрун.: Биохимия, 1964, 29, с.653-661; Скулачев В.П., Маслов С.П. и др. Холодовое разобщение окисления и фосфорилирования в мышцах белых мышей. / В жрун.: Биохимия, 1965, 28, с.70-79; Скулачев В.П. Аккумуляция энергии в клетке. М.: Наука, 1969; Скулачев В.П. Трансформация энергии в биомембранах. М.: Наука, 1972, 203 с.; Скулачев В.П. Гипотеза о биологической роли Na-К-градиента как резервной формы энергии. / Успехи современной биологии, 1978, 86, 3 (6), с.359-372; Скулачев В.П. Биохимия мембран./ Биоэнергетика.// Мембранные преобразователи энергии. М.: Высшая школа, 1989, 270 с.
[40] Коган А.Б., Косицкий Г.И., Кураев Г.А., Чораян Физиология человека и животных. М.: Высшая школа, 1984, 1 -2, 360 с., 287 с.
[41] Кондрашева М.Н. Трансаминазный цикл окисления субстратов в митохондриях, как единственный механизм адаптации к гипоксии (Тез. док. 1 Всесоюзн. конф.) Фармакологическая коррекция гипоксических состояний. 27 -28 янв. 1988г., с.68-69.
[42] Лукьянова Л.Д. Доклад на 1 Всесоюзн. конф. (Фармакологическая коррекция гипоксических состояний) 27-28 янв. 1988г., М. 1988г., с.107.
[43] Пасечник В.И. Электрострикционные измерения вязкоупругих свойств бислойных липидных мембран./ итоги науки и техники.// Ионные каналы и их модели./// М.: ВИНИТИ, 1982, с.267-307
[44] Nayler W.G., Yepez C.E., Fassold E. et al. Prolonged effect of propranolol on hypoxic heart muscle. / In.: Amer. J. Cardiology, 1978 y., 42, p.217.
[45] Trautwein W., Taniguchi J., Noma A. The effect of intracellular cyclic nucleotides and calcium on the action potential and acetylcholine response of isolated cardiac cells./In: Pflugers. Arch., 1982, 392, N 4, p.307-314
[46]Дементьева И.И., Бабалян Г.В., Соловьев Л.Е. Влияние комбинированной холодовой и фармакологической защиты на метаболизм миокарда при временной аноксии сердца. / В жрун.: Cor et Vasa, 1982, 24, 1, с.16-26
[47] Braunwald E., Maroko P. The reduction of infarct size an idea whose time (for testing) has come. / In.: Circulation, 1974 y., 50, 2, p.206-209.
[48] Blanco G., Adam A. et al. Complete homotransplantation of canine heart and lungs./In: Arch. Surgery, 1958, 76, 1, p.20-23
[49] Boyden P.A., Cranefield P.F., Gadsby D.C. Noradrenaline huperpolarizes cells of canine coronary sinus by increasing their permeability to potassium ions./In: J.Physiology Gr.Brit., 1983, 339, p.185-206
[50] Мачулин А.В. Консервация и реперфузия изолированного сердца в эксперименте. Автореф. дис. ... канд. мед. наук. М.: 1982, 31 с.
[51] Портной В.Ф. Подводные камни кардиоплегии. / В кн.: Хирургия сердца и сосудов. М.: 1978, с.144-145
[52] Бураковский В.И., Бокерия Л.А. Общие вопросы сердечно-сосудистой хирургии./ В кн.: Сердечно-сосудистая хирургия// Под ред. Бураковского В.И., Бокерия Л.А.// М.: Медицина, 1989, с. 11-44
[53] Малашенков А.И. Защита миокарда при операциях на аортальном клапане./ В жрун.: Вестник АМН СССР, 1981, 10, с.9-15
[54] Цукерман Г.И., Семеновский М.Л., Дубровский В.С., Малашенков А.И., Лифляндский Д.Б., Артюхина Т.В. Интраоперационная защита миокарда с помощью фармакологической кардиоплегии и локальной гипотермии при протезировании клапанов сердца. / В жрун.: Грудная хирургия, 1979, 1, с.22-29
[55] Подробнее взаимосвязь эффектов хроноинотропии и реакций гомеостатической регуляции будет освещена в этой же главе немного позже.
[56] Белоярцев Ф.Ф. Медико-биологические аспекты применения эмульсий перфторуглеродов, Сборник научных трудов ИБФ РАН, 1983, 186 с.
[57] Saks V.A., Bobkov Y.., Struma E., Creatine Phosphate/ Biochemistry, pharmacology and clinical efficiency, Torino, 1987, 270 p.
[58] Nichols J.R., Gonzales N.C. Increase in myocardial cell cGMP concentration in pressure-induced myocardial hypertrophy./In: J. Mol. and Cell. Cardiology, 1982, 14, N 3, p.181-183
[59] Schwartz K., Bastie D., Bouveret P., Oliviero P., Alonso S., Buckingham M. Alfa-Skeletal muscle action mRNAs accumulated in hypertrophied adult rat hearts./In: Circ. Res., 1986, 59, N 5, p. 551-555
[60] Хлоповская Е.И. Действие альфа-токоферола, селенита натрия, их комбинации и трентала на энергетический обмен, синтез РНК и белка при ишемии миокарда. Автореф. дис. ... канд.мед.наук. М.: 1980, 24 с.
[61] Salzmann J.L., Michel J.B., Bruneval P., Nlom M.O., Barres D.R., Camilleri J.P. Automated image analysis of myocardial collagen pattern in pressure and volume overload in rat cardiac hypertrophy./In: Anal. and Quant. Cytol. and Histol., 1986, 8, N 4, p.326-332
[62] Богач П.Г., Курский М.Д., Кучеренко Н.Е., Рыбальченко В.К. Структура и функции биологических мембран./ Киев: Вища школа, 1981, 335 с.
[63] Онищенко Н.А., Артамонов С.Д., Данилов М.А., Маевский Е.И., Лубяко А.А., Вазагашвили М.О., Илгавичуте Я. С., Жукаускас Г.Ю., Сафонов А.А. Энергетика процессов формирования резистентности органов к ишемии фармакологическими средствами./ Материалы Всесоюзной школы-семинара// Научные методы и технические средства защиты ишимизированных тканей. Устинов: 1986, с.8-10
[64] Лубяко А.А. Механизмы формирования и принципы диагностики противоишемической резистентности миокарда. Дис. ... Докт. биол. наук, М.: 1994, 487 с.
[65] Шумаков В.И. Механизмы адаптации трансплантированных органов. В жрун: Вестник РАН, 1996, том 66, N 12, с.1072-1078
[66] Jchevich N.V., Bidzilia J.P. Effect of physical exercise on changes of the hemodinamics in dogs with a denervated heart./In: Agressoloqie, 1975, 16, num.spec. B., p.49-53; Kammermeier H., Schmidt P., Jungling E. Free energy change of ATP-hydrolysis: a consul factor of early hypoxic failure of the myocardium?/In: J. Mol. and Cell. Cardiology, 1982, 14, N 5, p.267-277; Kaye M.P., Randall W.C., Hageman G.R., Geis W.P. Priola D.V. Chronology and mode of reinervation of the surgically denervated canine heart: functional and chemical correlates./In: Amer. J. Physiology, 1977, 233, N 4, H431-H437
[67] Jchevich N.V., Bidzilia J.P. Effect of physical exercise on changes of the hemodinamics in dogs with a denervated heart./In: Agressoloqie, 1975, 16, num.spec. B., p.49-53
[68] Kaye M.P., Randall W.C., Hageman G.R., Geis W.P. Priola D.V. Chronology and mode of reinervation of the surgically denervated canine heart: functional and chemical correlates./In: Amer. J. Physiology, 1977, 233, N 4, H431-H437
[69] Dausset J. (Ж.Доссе) Иммуногематология. М.: Медгиз, 1959 г., 637 с.
[70] Каштылян Л.С. Кумулятивная активность изофермента КФК-МВ у больных после хирургической коррекции врожденных пороков сердца. М.: 1984, 19 с.
[71] Сапрыгин Д.Б. Ферментативная диагностика и оценка повреждения миокарда у больных, оперированных на открытом сердце. Автореф. дис. ... докт. мед. наук. М.: 1986, 48 с.
[72] Лубяко А.А. Механизмы формирования и принципы диагностики противоишемической резистентности миокарда. Дис. ... докт. биол. наук, М.: 1994, 487 с.
[73] Возвращаясь к нашему конкретному примеру кардиомиоцита, в данном контексте, необходимо обязательно учитывать, что в условиях фармако-холодовой кардиоплегии система регуляции клеточного гомеостаза редуцирована до уровня внутриклеточной регуляции и подчинена таким образом законам функционирования изолированной клетки - конечного «исполнителя» физиологического ответа.
[75] Важно заметить, что ход приведённых последовательных рассуждений может быть экстраполирован на отношения любых участников гомеостатического ряда вне зависимости от уровня их расположения, ибо представляет по своей сути закон сохранения вещества, энергии и информации, приложенный к балансу отношений.
[76] Хотя, быть может, это лишь терминологическая погрешность изложения, связанная с толкованием категорий «неспецифическая». Мы трактуем от патофизиологического: «неспецифическая - есть типическая»
[77] как назвал их профессор Н.А. Фёдоров и его коллеги (Фёдоров Н.А. Биологическое и клиническое значение циклических нуклеотидов. М.: Медицина, 1979, 184 с.; Фёдоров Н.А., Радуловацкий М.Г., Чехович Г.Е. Циклические нуклеотиды и их аналоги в медицине. М, Медицина, 1990, 192 с.), где messenger – в переводе с английского - вестник, посыльный
[78] Если рассматривать, как пример, фармако-холодовую кардиоплегию с позиций патологической физиологии, то её можно отнести к одной из разновидностей гибернации (или гипобиоза), т.е. к консервации сердца, связанной с искусственным прекращением кровоснабжения, отменой механической и электрической активности сердца, децентрализации, насильственной отменой правил гомойтермии (322). При этом, факторы ишемии, отмены сокращений и децентрализации являются несомненно стрессовыми для сердца как органа, миокарда как ткани и кардиомиоцитов как клеток этой ткани. В то же время такая искусственная изоляция (без механического повреждения целостности структур) ставит орган, его ткани и клетки в очень сложные условия одновременного действия дефицита кислорода, не поступления субстрата, отсутствия гуморальной регуляции и химической блокады иннервации. Иными словами, миокард попадает в состояние тотальной ишемии и, одновременно, в условия резко ограниченных ауторегуляторных возможностей, редуцированных до уровня внутриклеточной регуляции.
[79] Это нам кажется вполне корректным, т.к. рассматриваемые в качестве примера, образца и представителя гомеостаза клеточного уровня, кардиомиоциты, в связи с «редукцией» систем их гомеостатической регуляции и сохранности морфологической организации, попавших в условия ишемии и (или) фармакохолодовой кардиоплегии, резонно рассматривать не как единую ткань, а как сумму структурно организованных, но регуляторно разобщенных клеток.
[80] Меерсон Ф.З. Теория индивидуальной адаптации к среде и профилактика стрессорных повреждений сердца. /Текст лекций// М.: УДН, 1981, 46 с.; Меерсон Ф.З. Адаптация, стресс, профилактика. М. Медицина, 1981г., с.278
[81] Кирпатовский В.И. Противоишемическая защита донорских почек. Дис. ... канд.мед. наук. М.: 1979, 190 с.
[82] Условно изолированной от окружающей клетку среды – от организма
[83] настоящим, действительно существующем (Артамонов С.Д., Онищенко Н.А. Использование метода дифференциальной калориметрии для изучения энергетических механизмов защитного действия противоишемических средств./В журн.: Фармакология и токсикология, 1983, 6, с.88-91; Артамонов С.Д. Динамика теплопродукции тотально ишемизированного изолированного сердца и связь ее изменения под действием витамина Е с полноценностью восстановления восстановления функции миокарда при его реоксигенации. Дис. ... канд. мед. наук М: 1986, 180 с).
[84] Лубяко А.А. Механизмы формирования и принципы диагностики противоишемической резистентности миокарда. Дис. ... Докт. биол. наук, М.: 1994, 487 с.
[85] Down W.H., Chasseand L.F., Ballard S.A. The effect of intravenously administered phosphocreatine on ATP and CP concentrations in the cardiac muscle of the rat./In: Arzneimittel Forsch/Drug Res. 1983, 39, p.552-554
[86] Doring H.J., Hauf G. Kontraktilitat sowie ATP- und kreatinphosphat-konzentrationen des meerscheinchenmyokards bei normoxie und verschiedenen hypoterenol und kalziumchlorid./In: Herz Kreislauf, 1977, 9, N15, p.926 -933
[87] Kammermeier H., Schmidt P., Jungling E. Free energy change of ATP-hydrolysis: a consul factor of early hypoxic failure of the myocardium?/In: J. Mol. and Cell. Cardiology, 1982, 14, N 5, p.267-277
[88] то, что часто связывают с дефицитом энергии
[89] Haber R.S., Loeb J.N. Early enticement of passive potassium efflux from rat liver by thyroid hormone: relation induction of Na-K-ATPase./In: Endocrinology, 1984, v.115, N 1, p.291-297
[90] Lullmann H. Relationship between Na-K-ATPase activity and electromechanical coupling in cardiac tissue./In: Progr. Pharmacol., 1978, 2, N 1, p.7-9
[91] Rayson B.M., Lowther S.O. Increased (Na+K+)ATPase activities in the hearts of hypertensive rats./In: J. Mol. and Cell. Cardiology, 1981, 13, Supple. N 3, p.6
[92] Lamers J.M., Stinis J.T., De Jonge H.R. On the role of cyclic AMP and Ca++-calmodulin-dependent phosphorylation in the control of Ca++-Mg++-ATPase of cardiac sarcolemma./In: FEBS Lett., 1981, 127, p.139-143
[93] Morcos N.,Ch., Drumond G.J. (Ca++Mg++)ATPase in enriched sarcolemma from dog beast./In: Biochim., Biophys. Acta, 1980, 598, p.27-39; Morcos N.Ch. Isolation of frog Heart sarcolemma possessing (Ca++-Mg++)ATPase and Ca++ pump activities./In: Biochim. Biophys. Acta., 1981, v.643, p.55-62; Morcos N.Ch. Localisation of (Ca++Mg++)ATPase, Ca++ pump and other ATPase Activities in cardiac sarcolemma./In: Biochim. and Biophys. acta, 1982, 688, N 3, p.747-756
[94] Panagia V., Lamers J., Singal P., Dhalla N. Ca++ and Mg++ dependent ATPase activities in the deoxycholate-treated rat heart sarcolemma./In: Int. J. Biochem., 1982, 14, N 5, p.387-397
[95] Caroni P., Carafoli E. The Ca-pumping ATPase of heart sarcolemma, characterisation, calmodulin dependence and partial purification./In: J. Biol. Chem. 1981, 256, p.3263-3270
[96] Федоров Н.А. Биологическое и клиническое значение циклических нуклеотидов. М.: Медицина, 1979, 184 с.
[97] Mistry G., Drummond G.I. Heartmicrovessels: presents of adenylate cyclase stimulated by catecholamines, prostoglandins and adenosine./In: Microvasc. Res., 1983, 26, N 2, p.157-169
[98] Лубяко А.А. Регуляция клеточного гомеостаза (На примере кардиомиоцита). Л.: 1990, 66 с; Лубяко А.А. Механизмы формирования и принципы диагностики противоишемической резистентности миокарда. Дис. ... докт. биол.наук, М.: 1994, 487 с.
[99] а также кардиоплегических растворов при операциях на сердце и иных консервирующих растворах при экстракорпоральном сохранении органов, тканей, клеток, пересадках почек и печени, реплантациях конечностей
[100] Kotic A., Yanachek K. (А.Котык, К.Яначек) Мембранный транспорт. М.: Мир, 1980 г., 338 с.
[101] Keely S.L., Corbin J.D. Involvemen of cAMP-dependent protein kinase in the regulation of heart contractile force./In: Amer. J. Physiology, 1977, 233, N 2, H269-H275
[102] Кukovetz W. Poch G. Pharmakologie der betablokators./In: Adv. Clinical Pharmacol. 1976., v. 11, p.1189-1191.
[103] Бреслер В.М., Никифоров А.А. Транспорт органических кислот через плазматические мембраны. Л.: Наука, 1981, 202 с.
[104] Hochachka P.W., Somero G.N. (П.Хочачка, Дж.Сомеро) Стратегия биохимической адаптации. М.: Мир, 1977 г., 398 с.; Hochachka P.W., Somero G.N. (П.Хочачка, Дж.Сомеро) Биохимическая адаптация. М.: Мир, 1988, 567 с.
[105] Костюк П.Г. Кальций и клеточная возбудимость. М.: Наука, 1086, 223 с.
[106] Libbert E. (Э.Либберт) Основы общей биологии. М.: Мир, 1982 г., 412 с.
[107] Меерсон Ф.З., Брегер А.М. Сопряжение возбуждения с сокращением и расслаблением сердечной мышцы. Ижевск: 1976, 91 с.
[108] Kokuban Sh., Nishimura M., Noma A., Irisawa H. Membrane currents in the rabbit atrioventricular node cell./In: Pflugers Arch., 1982, 393, N 1, p.15-22
[109] Mc Donald T.F., Mc Leod D.P. Anoxia-Recovery Cycle in ventricular muscle: Action potential duration, contractility and ATP content./In: Pflugers Arch.ges. Physiology, 1975, 325, p.305-322; Mc Donald T.F. The slow inward calcium current in the heart./ In.: Annu. Rev. Physiology, Palo Alto, Calif., 1982, v.44, p.425-434; Mc Donald T.F., Электромеханическое сопряжение. Связь медленного входящего тока с сокращением./В кн.: Физиология и патофизиология сердца. М.: Медицина, 1988, т.1, с.278-295
[110] Chapman R.A., Coray A., Mc Guigan J.A. Sodium/calcium exchange in mammalian ventricular muscle. / In.: J. Physiology, 1981, 318, P13-P14; Chapman R.A., Fozzard H.A., Friedlander I.R., January C.T. Effects of Ca++Mg++ removal on a[Na]i, a[K]i and tension in cardiac Purkinje./In: Amer. J. Physiology, 1986, 251, 6, Pt.1, p. 920-927
[111] Chapman R.A., Coray A., Mc Guigan J.A. Sodium/calcium exchange in mammalian ventricular muscle. / In.: J. Physiology, 1981, 318, P13-P14; Chapman R.A., Fozzard H.A., Friedlander I.R., January C.T. Effects of Ca++/Mg++ removal on a[Na]i, a[K]i and tension in cardiac Purkinje./In: Amer. J. Physiology, 1986, 251, 6, Pt.1, p. 920-927
[112] Так известно, что катехоламины, норадреналин, в частности, вызывают состояние гиперполяризации мембран кардиомиоцитов (239, 261, 353). А это, уже факт существования прямой связи активности аденилатциклазы и поляризации мембран. Но и это еще не достаточно убедительно. По данным P.A.Boyden et al. (1983), увеличение концентрации калия в перфузате[112] способно вызвать значительное снижение эффекта гиперполяризации кардиомиоцитов[112], вызванной норадреналином (239). Наличие же такой «инвертированной» связи: во-первых, утверждает ее существование, а во-вторых, не опровергает и существование прямой: гиперполяризация есть посылка для торможения высвобождения катехоламинов, деполяризация - стимул.
[113] Каменская М.А. Нейротоксины в физиологических исследованиях./В кн.: Итоги науки и техники.// Физиология человека и животных. М.: ВИНИТИ, 1982, 26, 104 с.
[114] Так известно, что катехоламины, норадреналин, в частности, вызывают состояние гиперполяризации мембран кардиомиоцитов (239, 261, 353). А это, уже факт существования прямой связи активности аденилатциклазы и поляризации мембран. Но и это еще не достаточно убедительно. По данным P.A.Boyden et al. (1983), увеличение концентрации калия в перфузате[114] способно вызвать значительное снижение эффекта гиперполяризации кардиомиоцитов[114], вызванной норадреналином (239). Наличие же такой «инвертированной» связи: во-первых, утверждает ее существование, а во-вторых, не опровергает и существование прямой: гиперполяризация есть посылка для торможения высвобождения катехоламинов, деполяризация - стимул.
[115] Пасечник В.И. Электрострикционные измерения вязкоупругих свойств бислойных липидных мембран./ итоги науки и техники.// Ионные каналы и их модели./// М.: ВИНИТИ, 1982, с.267-307
[116] Лайтфут Э. Явления переноса в живых системах. М.: Мир, 1977, 518 с.; Богач П.Г., Курский М.Д., Кучеренко Н.Е., Рыбальченко В.К. Структура и функции биологических мембран./ Киев: Вища школа, 1981, 335 с.; Kotic A., Yanachek K. (А.Котык, К.Яначек) Мембранный транспорт. М.: Мир, 1980 г., 338 с.
[117] Пасечник В.И. Электрострикционные измерения вязкоупругих свойств бислойных липидных мембран./ итоги науки и техники.// Ионные каналы и их модели./// М.: ВИНИТИ, 1982, с.267-307; Богач П.Г., Курский М.Д., Кучеренко Н.Е., Рыбальченко В.К. Структура и функции биологических мембран./ Киев: Вища школа, 1981, 335 с.; Kotic A., Yanachek K. (А.Котык, К.Яначек) Мембранный транспорт. М.: Мир, 1980 г., 338 с.
[118] это предположение, имеющее косвенные подтверждения
[119] Лайтфут Э. Явления переноса в живых системах. М.: Мир, 1977, 518 с.; Kotic A., Yanachek K. (А.Котык, К.Яначек) Мембранный транспорт. М.: Мир, 1980 г., 338 с.; Богач П.Г., Курский М.Д., Кучеренко Н.Е., Рыбальченко В.К. Структура и функции биологических мембран./ Киев: Вища школа, 1981, 335 с.
[120] Ивков В.Г., Берестовский Г.Н. Липидный бислой биологических мембран. М.:Наука, 1982, 224 с.
[121] Это очень важный момент, ибо составляет суть практического примера, доказывающего прямую связь и, следовательно, механизм трансформации энергии и информации волнового свойства в материальное на молекулярном уровне. А это, в свою очередь, один из ключевых и исторически первичных механизмов централизованного и оперативного управления со стороны Высшего гомеостаза, наряду с системами управления кластерного типа, фотосинтезом и т.д. Об этом подробно см. в разделе 3.
[122] Меерсон Ф.З., Брегер А.М. Сопряжение возбуждения с сокращением и расслаблением сердечной мышцы. Ижевск: 1976, 91 с.; Kotic A., Yanachek K. (А. Котык, К. Яначек) Мембранный транспорт. М.: Мир, 1980 г., 338 с.; Lamers J.M., Stinis J.T., De Jonge H.R. On the role of cyclic AMP and Ca++-calmodulin-dependent phosphorylation in the control of Ca++-Mg++-ATPase of cardiac sarcolemma./In: FEBS Lett., 1981, 127, p.139-143
[123] Пример одного из следствий в механизме управления кластерного типа при формировании энергетически выгодных неравновесий.
[124] в соответствии с классификацией Ф.З.Меерсона
[125] по той же классификации Ф.З.Меерсона
[126] Этот момент представляется чрезвычайно своевременным для очень важной ремарки: поскольку клетка – это очень древний уровень в организации материи, фактически – первый представитель «живого» на Земле, то именно в ней, в её древних, диалектически детерминированных реакциях акцепта информации, наиболее полно отражены исторические связи, организующие всепроникающую систему центрального управления процессом термодинамически выгодного самоусовершенствования и самоусложнения, т.е. рождения энтропии (по И.Р. Пригожину)
[127] Физиологический же ответ, как мы уже неоднократно указывали – есть суть, составляющая механизм «принципа обратной связи И.И.Шмальгаузена» в общей «теории информации и информационных пространств».
[128] Сафронова В.Г. Исследование роли быстрого калиевого тока и нейрогуморальных факторов в регуляции электрогенеза и сократимости миокарда. Аватореф. дис. ... канд. биол. наук, Пущино-на-Оке, 1982, 16 с.
[129] Nayler W.G., Poole-Wilson A. Hypoxia and Ca++ metabolism. / In.: J. Mol. and Cell. Cardiology, 1979 y., 11, 7, p.683-706.; Nayler W.G., Crinwald P.M. Dissociation of Ca++ accumulation from protein release in calcium paradox: effect of barium. / In.: Amer. J. Physiology, 1982, 242, N 2, H203-H210.
[130] Кокоз Ю.М., Кобринский Е.М., Фрейдин А.А., Лазарев А.В. Активация потенциал зависимого Са++ тока в сердечных волокнах алкалоидом форсколином. Препринт. Пущино-на-Оке, 1985, 14 с.
[131] Ochi R. Role of divalent cations in slow responses./In: J. Mol. and Cell. Cardiology, 1977, 9, N 11, Suppl., p.17; Ochi R. Inhibition of Ca-dependent slow inward current by acetylcholine in mammalian ventricular muscle. / In.: J. Mol. and Cell. Cardiology, 1981 y., 13, 2, p.123.
[132] Бабский Е.Б., Богданова Е.В. Динамика энергетического обмена в миокарде в течение сердечного цикла./ В кн.: Метаболизм миокарда. М.: 1975, с.260-267
[133] Chapman R.A., Coray A., Mc Guigan J.A. Sodium/calcium exchange in mammalian ventricular muscle. / In.: J. Physiology, 1981, 318, P13-P14; Chapman R.A., Fozzard H.A., Friedlander I.R., January C.T. Effects of Ca++-Mg++ removal on a[Na]i, a[K]i and tension in cardiac Purkinje./In: Amer. J. Physiology, 1986, 251, 6, Pt.1, p. 920-927
[134] падение гликогена, молочной кислоты, сначала падение, а затем резкая активация ЛДГ, рост ПВК
[135] преобразование специфической информации в доступную для «понимания» или неспецифическую
[136] Мархасин В.С., Мильштейн Г.Н. Моделирование влияния ритма на силу сокращения сердечной мышцы./ В журн.: Биофизика, 1978, ХХШ, 4, с.674-681; Мархасин В.С. Механизмы нарушения сократительной функции миокарда при хронической сердечной недостаточности. Автореф. дис. ...докт.биол.наук. Купавна: 1983, 39 с.
[137] Изаков В.Я., Иткин Г.П., Мархасин В.С., Штенгольд Е.Ш., Шумаков В.И., Ясников Г.П. Биомеханика сердечной мышцы. М.: Наука, 1981, 325 с; Изаков В.Я., Антьюфьев В.Ф., Проценко Ю.Л. Ритмоинотропные явления в сердце человека./ В жрун.: Физиология человека. 1983, 9, 5, с.740-747; Изаков В.Я. Исследование клеточных систем регуляции механической активности миокарда (биомеханический аспект). Автореф. дис.... докт.мед.наук, Томск, 1985, 43 с.
[138] t – зависимые (зависимые от времени)
[139] Лубяко А.А., Эстрин Е.С., Александров К.Ю., Пустовалов П.Ф. Способ исследования функционального состояния сердца./Заявка на авт. свидетельство N 4855092/14, 83699 от 31 июля 1990 г.; Лубяко А.А., Курапеев И.С. Энергобаланс сердечно-сосудистой системы во время искусственного кровообращения./В жрун: Грудная и сердечно-сосудистая хирургия, 1994, No5, с.34-36; Лубяко А.А. Механизмы формирования и принципы диагностики противоишемической резистентности миокарда. Дис. ... Докт. биол. наук, М.: 1994, 487 с.
[140] Лубяко А.А., Кирпатовский В.И. Значение адаптационных реакций полоски миокарда при изменении режимов электростимуляции для оценки энергосостояния сердца крысы. / В сб. трудов института// Трансплантация и искусственные органы. М.: 1981, с.107-110
[141] часть структур «отдыхает» пока друга часть, готовящаяся их заменить, работает
[142] Только в небольшом количестве работ подробно рассматриваются механизмы расслабления (154, 205, 231, 333) и сопряжение их с процессом сокращения (154)
[143] при неизменной длине волокна
[144] при неизменном объеме
[145] Дорофеев Г.И., Кожемякин Л.А., Ивашкин В.Т. Циклические нуклеотиды и адаптация организма. Л.: 1978
[146] Inui M., Tada M., Kadoma M., Yamada M., Kuzuya T. Effect of cyclic AMP and calmodulin dependent phosphorylation of phospholamban on calcium transport ATP-ase of cardiac sarcoplasmatic reticulum. / In.: J. Mol. and Cell. Cardiology, 1981, 13, Suppl. N 2, p.85.
[147] Seuss H. Untersuchunqen uber die Bedutung von Calcium und Magnesium fur die Kontraktion des Herxmuscels. / In.: Diss. Dokt. Naturwiss, Fak. Chem. Biol. und Geowiss. Techn. Univ. Munchen, 1980, 51 S.
[148] Goto M., Urata M., Hyodo T. Na-Ca exchange tension and membrane current examined on the bull forge myocardium under voltage damp. / In.: J. Mol. and Cell. Cardiology, 1981, 13, Suppl. 2, p.24.; Ленинджер А.Л. Основы биохимии. М.Мир, 1985г., 1057 с.; Kotic A., Yanachek K. (А.Котык, К.Яначек) Мембранный транспорт. М.: Мир, 1980 г., 338 с.
[149] которую можно уверенно назвать основным механизмом поддержания постоянным соотношения связанного и свободного кальция (как, впрочем, и любого химического вещества, способного к диссоциации)
[150] в соответствии с классификацией Ф.З.Меерсона
[151] Гомеостаз органоидов и органелл носит больше условный характер ввиду его исторически рудиментарного происхождения и абсолютным отсутствием изначальных механизмов для самостоятельности и независимости в поддержании собственного гомеостаза.
[152] который был продемонстрирован на примере кардиомиоцитов
[153] Т.е. является диагностически значимым для системной диагностики
[154] если эта система сохранна, а информация не избыточна
[155] на тех же условиях
[156] для кардиомиоцитов, - по реакциям экстра- и постэкстрасистолической потенциации, как естественной, циклически повторяющейся «нагрузочной пробе». При этом, если реакция «возмущения» сопровождается для увеличением тонического напряжения (напряжения покоя), связанного только с развивающимся дефицитом АТР (скорости синтеза АТР), то завершающая процесс срочной адаптации фаза должна быть фазой расслабления, т.к. она связана с ликвидацией такого дефицита и т.д.
Оглавление Введение Часть 1 Часть 2 Часть 3 Заключение
Глава 1 Глава 2 Глава 3 Глава 4 Глава 5 Глава 6 Глава 7 Литература