Оглавление Введение Часть 1 Часть 2 Часть 3 Заключение
Глава 1 Глава 2 Глава 3 Глава 4 Глава 5 Литература
Глава 3
Основополагающие учения
«Идеи единого гомеостатического устройства»
«…принципы защиты на уровне клетки в основном приложимы и к человеку,
и даже к целым сообществам людей»
Ганс Селье
И так, что же это: «парабиоз», «физиологическая лабильность», «доминанта», «функциональная система»? Что такое «адаптационный синдром»? Что есть «эволюция»?
Наверное, лучше всего на эти вопросы смогли бы ответить авторы этих понятий, - люди, кто непосредственно ввел эту терминологию в устоявшийся и такой сложный образ мышления современников.
Однако, прежде чем приступить к изложению содержания этой главы и ответить на поставленные вопросы, хотелось бы акцентировать внимание коллег на двух особенностях в её содержании:
1. Во-первых, здесь весьма часто будут приведены дословные цитаты авторов, характеризующих то или иное понятие, те или иные суждения о них. Мы умышленно использовали этот прием, исходя из нескольких весьма принципиальных для нас соображений:
· Мы полагали облегчить коллегам возможность восстановить в памяти подлинные мысли авторов, их комментарии, которые в процессе частого употребления и толкования подчас приобретают иные оттенки, а иногда, - вовсе другой смысл.
· Нам хотелось, чтобы коллеги имели удобную возможность оценить правомочность и корректность нашего понимания и толкований при использовании общеизвестных академических учений классиков, положенных в основу формируемой идеи;
· Мы не считали себя вправе, опираясь на авторитет основоположников целого ряда современных дисциплин и наук, являющихся фактическими авторами, пусть разрозненных, но частей самостоятельного учения, коей мы видим «Идею единого гомеостатического устройства», толковать их высказывания и мысли на этом этапе изложения;
· Не хотелось бы также предлагать хоть какой-то повод слишком строгим критикам и рецензентам говорить о изменении смысла «вырванного из авторского текста» толкования того или иного процесса, явления или просто мысли, теряя на этом их и своё время в бесцельных объяснениях.
2. Во-вторых, в воспроизводимых цитатах речь идет, в основном, об организме, органах, тканях, клетках человека и животных, биосфере или популяции. Это связано, прежде всего, с тем, что общей «Идеи единого гомеостатического устройства» не только мира, но и биосферы пока нет. Она только «складывается», воссоздается заново, а названные участники гомеостатических отношений не только доступны для исследования, но и в достаточной мере хорошо изучены. Это тем более обнадеживает, ибо из пяти названных ранее уровней гомеостаза, как мы уже отмечали, выделенных в самостоятельные иерархические уровни (органелла, клетка, организм, планетарная биосфера, метагалактика), четыре являются доступными для исследования.
И хотя общей теории о «едином гомеостатическом устройстве» пока не существует даже на уровне самостоятельного и, в достаточной мере, современного «учения о гомеостазе», но разобщенных материалов очень и очень много. Их более чем достаточно для внимательного анализа и объединения. Но они разрознены, часто противоречивы и, что называется, «не когерентны». Тем не менее, самостоятельность предлагаемой идеи формируется этой книгой, хотя подготовка к её написанию осуществлялась в течение более пятнадцати лет.
Отсутствие объединяющего теоретического начала, о котором мы говорим, можно будет отчетливо почувствовать и в приводимых авторских цитатах и в комментариях к ним. Это ещё один повод и ещё один не маловажный аргумент в пользу так называемого «прямого цитирования» основоположников.
В связи с этим, нам кажется извинительным стремление максимально объективизировать и саму идею, и её предпосылки, представив мысли и рассуждения о них «авторскому исполнению».
Ведь, в конечном итоге, эта книга являет собой «материализованное стремление» объединения, систематизации и экстраполяции знаний о частном на общее и «очень большое» Всеобщее, выносимое нами на первый суд научной общественности, а потому хотелось бы создать максимально открытую атмосферу для объективности и, по возможности, доброжелательности суждений.
Этой книгой мы хотели бы также показать действительное место «учения о гомеостазе» и «Идеи единого гомеостатического устройства мира» для биологии, естествознания, философии и, конечно, - нормальной и патологической физиологии, как основополагающих дисциплин медицинской науки, кои являются главным и основным объектом наших профессиональных интересов.
Основные положения эволюционной теории Чарльза Роберта Дарвина (1809-1882) «Происхождение видов путём естественного отбора» (1859)
Сам Чарльз Дарвин особенно подчёркивал основополагающую роль научных работ своих предшественников Ж.Л. Бюффона (1707-1788), Ж.Б. Ламарка (1744-1829), Г.И. Менделя (1822-1884), подготовивших весьма серьезную идеологическую базу для рождения и обоснования эволюционной теории. Уже после выхода в свет его работы «Происхождение видов путем естественного отбора» в 1859 году, «эволюционная теория» и «дарвинизм» стали отождествляться.
Наверное, не стоило бы даже останавливаться на столь грандиозном и практически беспрекословно принятом учении об эволюции живого на Земле, если бы оно не являлось, по существу, одной из самых ярких иллюстраций адаптивно-приспособительных возможностей для самого широкого спектра представителей (видов) «живой материи», вершиной творения которой стал организм высших животных и человека.
Нам важен именно этот факт, ибо гомеостаз биосферы и гомеостаз организмов формировался и продолжает формироваться здесь на Земле по общим законам развития и бытия вне зависимости от нашей воли и сознания.
Может показаться, что мы, в попытке обобщения, невольно вынуждены уходить от частностей. Таким понятием, как «гомеостаз организма высших животных и человека» мы, как бы «отсекаем» весь яркий и разнообразнейший мир живого, концентрируя внимание только на одном, «более интересном» для нас объекте. Это вовсе не так.
Мы действительно не имеем ни намерений, ни желания превращать наше изложение в излишне детализованное «чудовище», наполненное частными примерами гомеостаза от многообразия видов и форм. В этом нет необходимости. Мы пытаемся рассмотреть общее и всеобщее в многообразии частного и потому значение таких обобщающих работ как учение Чарльза Дарвина просто неоценимо вместе со своими «спорными» и «бесспорными» постулатами.
Гомеостаз - везде гомеостаз. Везде и для всех видов живого мира органелла – это внутриклеточное содержимое, отделенное от внешней среды мембраной, внутренняя среда которой отлична от окружающей. Для органеллы - это «гомеостаз органеллы». Для клетки – это «гомеостаз клетки». Для организма – это «гомеостаз организма». Кто бы ни «владел» этим морфофункциональным материалом, этим «кирпичиком» мироздания, гомеостаз есть гомеостаз. Микроорганизмы ли это, амфибии ли, черви, рыбы, земноводные, птицы, млекопитающие, человек, биосфера или Вселенная. Из одних и тех же «кирпичиков» может быть построен храм, а может быть сооружён дворец съездов. Только в первом случае это сооружение станет домом Господним, а во втором – местом официальных сборов избранных представителей искусственного гомеостаза.
И так, Ж.Л. Бюффон, Ж.Б. Ламарк и Г.И. Мендель показали:
· Все органические формы не представляют чего-то навсегда установленного, неизменного, а меняются в процессе развития;
· Их изменчивость и развитие обусловлены наследованием признаков через совокупную чистоту гамет;
· Все существующие виды растений и животных являются результатом этого развития.
Чарльз Дарвин и независимо от него Альфред Рассел Уоллес в период 1823-1913 гг. обосновали принципы естественного отбора и сформировали чёткое представление о «борьбе за существование» как механизме этого отбора. В результате ими были выдвинуты следующие основные положения:
· «Близко родственные организмы (например, родители и дети) сходны между собой, но у них есть наследственные различия;
· Эти различия становятся более заметными, если рассматривать длинные ряды предков и потомков;
· Разные признаки изменяются с разной быстротой, так что одни признаки могут быть более давним филогенетическим приобретением, чем другие;
· Производится больше потомков, чем может выжить, и потому происходит отбор «наиболее приспособленных»;
· Основой естественного отбора является совместное и неразрывное действие трёх факторов: изменчивости, наследственности и перенаселённости, существующей в живой природе;
· Эволюция живой материи… необратима», (но повторяема). Конец цитаты.
Говоря о «перенаселённости» Чарльз Дарвин вплотную подошел к пониманию главного фактора, регулирующего численность вида того или иного представителя живого мира. Т.е. уже тогда им была фактически обозначена «популяция», как объект регулирования, а инструментом регулирования численности такой популяции практически был назван фактор плотности участников популяции на единице площади.
Говоря о «необратимости» эволюции и уровне её развития в сегодняшнем дне, за порогом «конца света», о котором можно было услышать на каждом официальном и неофициальном «углу», за пороге нового века и нового тысячелетия, хочется процитировать замечательные слова блистательного русского ученого и естествоиспытателя профессора Н.И. Вавилова:
В своем интервью Жану Аннри Моран, журналисту одной из французских газет, Николай Иванович Вавилов сказал: «Человек является биологической разновидностью, которая находится лишь в первой стадии своей истории. Она ещё сконцентрирована в своей колыбели... »[1]. Это во истину прекрасные, горькие и, скорее всего, пророческие слова во всех отношениях.
Биогенетический закон профессора Эрнста Геккеля (1834-1919) и профессора Фрица Мюллера (1821-1897) при использовании фактических данных профессора А.О. Ковалевского (1840-1901):
Вообще, хотя собственно термин «биогенетический закон» ввёл профессор Э. Геккель ещё в 1866 году, его проявления были замечены много раньше. Это были немецкий естествоиспытатель и натурфилософ Лоренц Окен (1779-1851) и русский профессор биологии Карл Францевич Рулье (1814-1858).
Используя натурфилософские построения Ф. Шеллинга и Г. Гегеля, профессор Л. Окен рассматривал всё многообразие живых организмов как результат развития и превращений, каждая ступень которого реализует некую «первичную форму» или «архетип», выдвинув гипотезу происхождения жизни из первичной слизи, образовавшейся в море, и гипотезу возникновения более сложных организмов из элементарных частиц.
Ещё до выхода в свет «Происхождения видов» (1859) Чарльза Дарвина, профессор К.Ф. Рулье указывал (1852) на опыт выведения новых пород животных и их акклиматизацию, как ключ к пониманию движущих сил эволюции в естественных условиях, подчёркивая, что наследственность определяется исторически сложившимися условиями, а изменчивость является процессом приспособления организма к условиям существования.
Между тем, авторство на биогенетический закон заслуженно принадлежит Эрнсту Геккелю и Францу Мюллеру. Закону, устанавливающему, что в течение своего индивидуального развития (в особенности – зародышевого) все без исключения живые существа повторяют главнейшие этапы развития всего ряда своих предковых форм.
В оригинальном же исполнении основные постулаты этого закона звучат следующим образом: «Онтогенез – есть краткое и быстрое повторение филогенеза, обусловленное физиологическими функциями наследственности (размножения) и приспособления (питания). Филогенез определяет стадии онтогенеза, а онтогенез не влияет на ход исторического развития».
Важнейшие факторы «Синтетической теории эволюции» профессора Т.Г. Хаксли (1825-1895) и доктора Дж. Симпсона (1878-1954):
В последарвиновский период, основным содержанием которого стал анализ механизмов и закономерностей эволюции, в разработку эволюционного учения внесли вклад многие учёные. Среди них: В.О. Ковалевский (1842-1883), Т. Морган (1866-1945), А.Н. Северцов (1866-1936), Н.И. Вавилов (1887-1943), Дж. Холдейн (1892-), Т.Г. Хаксли (1825-1895), Дж. Симпсон (1878-1954) и многие другие.
В развитие идей Ч.Дарвина, «синтетическая теория эволюции», сложившаяся заслугами Т.Г. Хаксли и Дж. Симпсона в 30-ых годах XX века, существенно дополнило объяснение существующего разнообразия видов и приспособляемости организмов как результат действия в основном двух факторов:
· «Непрерывного появления наследственных отклонений и отбора, осуществляемого факторами внешней среды, где онтогенез и физиологические процессы рассматриваются как реализация наследственной информации, а филогенез – как формирование и испытание всех старых и новых наследственных информационных программ;
· Способность организмов к самоорганизации, как самовоспроизводящихся систем, ограничивающих роль случайностей и происшедших, и происходящих путем творческих актов».
Принципы «изменчивости, наследственности и естественного отбора» профессора А.Н. Северцова (1866-1936), профессора К.А. Тимирязева (1843-1920), профессора В.И. Вернадского (1863-1945) и профессора Н.И. Вавилова (1887-1943):
С позиций современного эволюционного учения важнейшими факторами эволюции являются наследуемые мутации (изменчивость и наследственность) и естественный отбор.
«Изменчивость» - это свойство живых организмов существовать в различных формах. По механизмам возникновения, характеру изменений признаков различают несколько типов изменчивости:
1. Изменчивость наследственная или генотипическая (по Чарльзу Дарвину – «неопределённая изменчивость»), обусловленная возникновением новых генотипов, приводящая к изменению фенотипа в соответствии законам Г.И. Менделя. В её основе могут лежать мутации, и тогда она носит название – «мутационная изменчивость» или новые комбинации аллелей, образующиеся за счёт закономерного поведения хромосом в мейозе и при оплодотворении, или за счёт рекомбинаций. Тогда изменчивость получает название «комбинированная».
2. Изменчивость ненаследственная или модификационная (по Чарльзу Дарвину – «определённая изменчивость»), отражающая изменения фенотипа под действием условий существования организма, не затрагивающая генотип, хотя степень изменчивости может определяться и определяется именно генотипом («чистотой гамет» по Г.И. Менделю).
3. Изменчивость онтогенетическая, отражающая реализацию закономерных изменений в ходе индивидуального развития организма (морфогенез) или его клеток (дифференцирование). При этом типе изменчивости генотип остается неизменным, хотя изменчивость безусловно детерминирована генетическими факторами («чистотой гамет» по Г.И. Менделю).
4. Существуют ещё несколько вариантных имён изменчивостей:
-
«фенотипическая»;
-
«генотипическая»;
-
«альтернативная»,
-
«прерывистая» или «качественная»;
-
«флуктуирующая»,
-
«непрерывная» или «количественная».
Мы полагаем, что не стоит тратить время на комментарии к ним, т.к. в этой узко специализированной терминологической дискуссии пока нет предмета интересов для нашего изложения.
Для нас более важен факт того, что изменчивость, проявившаяся в процессе длительной «долговременной» (по Ф.З. Меерсону) адаптации к внешним условиям, способна к наследованию (мутационная изменчивость), если внешние условия или их колебания были так сильны, что эти изменения вызвали изменение органических структур организма и наследовались. Адаптация же носила при этом не только долговременный характер, но и позволила сформировать так называемый «системный структурный след» (Ф.З. Меерсон) – «генетическую память клетки».
Если же по каким-либо обстоятельствам произошедшие изменения не наследовались (модификационная изменчивость), но имели место быть у данной особи или популяции, то адаптационные процессы компенсировали избыточное действие факторов внешней среды без привлечения «генетической памяти», но при её участии. Либо факторы были слабы, либо особь и/или популяция оказались ослаблены и/или обречены на исчезновение.
«Наследственность» - это свойство организмов обеспечивать материальную и функциональную преемственность между поколениями.
«Наследственность», как это было сказано выше, может реализоваться в различных вариантах в зависимости от особенностей генотипа организма и условий внешней среды. Исторически возникло и развивалось представление о «наследственности» как отражении существования материальной субстанции, обеспечивающей сходство организмов в ряду поколений и многообразие их видов. В связи с этим, «наследственность» принято рассматривать как свойство, имеющее несколько типов:
1. «Хромосомная наследственность» - материальная преемственность между поколениями признаков, свойств, информации, локализованной в локусах и аллелях хромосом.
2. «Цитоплазматическая наследственность» - материальная преемственность между поколениями признаков, свойств, информации, локализованной в молекулах ДНК и РНК.
3. «Сигнальная наследственность» - материальная преемственность между поколениями, осуществляемая передачей информации в процессе обучения от одного поколения другому. Основа такого вида преемственности связана, прежде всего, с условно рефлекторной деятельностью высших животных и человека.
В рамках нашего изложения следует подчеркнуть, что «наследственность», а корректней было бы сказать: «наследуемость» или «наследование» признаков, свойств, информации есть не только главный фактор и базовый механизм эволюционного процесса. «Наследование» - это центральный аргумент «генетической памяти» адаптивных процессов в отношениях гомеостатической системы организмов с внешней для них средой. «Наследование» – это генетическая память мировой истории.
«Естественный отбор» - это совокупный движущий фактор в механизме эволюционной адаптации, рождённый в борьбе за существование с условиями внешней среды, плотностью популяции и межвидовых отношений. Фактор, постоянно производящий историческую базу для наследования наиболее устойчивых изменчивостей.
Таким образом, совокупность перечисленных факторов необходима и достаточна для характеристики осуществляемого во времени и пространстве эволюционного процесса, демонстрирующего высшую форму гомеостатической лабильности на уровне отношений участников планетарного гомеостаза: организм – биогеоценоз.
«Естественный отбор», непосредственно воздействуя на фенотипы организмов, вызывает наследование не отдельных признаков, свойств и аллелей, а целых генотипов, обладающих определенной «нормой реакции», о которой подробнее пойдёт речь ниже (Часть 2, глава 2). Сейчас важно, что регуляторной единицей эволюции гомеостатических систем, как постоянного и необратимого процесса долговременной адаптации, являются популяции.
Принцип профессора В.И. Вернадского (1863-1945) - единое геохимическое устройство «живого вещества» (1928):
Рассматривая вопросы формирования биосферы и ноосферы в процессе эволюции, профессор В.И. Вернадский писал: «В геохимическом аспекте, входя как часть в мало изменяющуюся, колеблющуюся около неизменного среднего состояния биосферу, жизнь, взятая как единое целое, представляется устойчивой и неизменной в геологическом времени.
В сложной организованности биосферы в пределах живого вещества происходят только перегруппировки химических элементов, а не коренные изменения их состава и количества». Конец цитаты.
Эти несложные, на первый взгляд, для понимания идиомы профессора В.И. Вернадского носят не только принципиально важный характер, они основополагают в своём предвидении ещё непознанное нами, но очень простое по существу, явление.
С одной стороны «необратимость» эволюционного процесса, как бы подразумевает обязательным факт движения, т.е. изменений, мутаций, модификаций; с другой – имеет фактическое «постоянство состава биосферы в геологическом времени».
Это не противоречие, это факт и факт изумительный, поскольку он указывает на то, что материя, как вечная субстанция, может быть и организована, и реорганизована в рамках единой компартментализованной гомеостатической системы. Это прямое указание всеобщего закона «отрицания отрицания» на необратимость движения в бесконечность рекомбинаций.
Таким образом, на один из вопросов, поставленных в начале этой главы: «Что есть эволюция?» с позиций предлагаемой «Идеи гомеостатического устройства» – мы уже можем ответить уже теперь: Эволюция - есть объективная реальность, отражающая необратимое и бесконечное движение в развитии и преобразованиях материи[2].
В плане нашего изложения, представляется вполне допустимым предложить «развернутую» формулировку, применимую как пояснение:
Эволюция – есть всеобщая закономерность (объективная реальность), отражающая необратимое движение в развитии и преобразовании материи, составляющая единую форму адаптационно-приспособительных реакций гомеостатических систем, выраженную в наследуемой изменчивости наиболее устойчивых признаков, свойств и информации в борьбе за выживаемость (сохранение собственного гомеостаза).
Иными словами эволюция – есть генеральная форма адаптации гомеостатических систем к изменениям внешней среды в обмене веществом, энергией и информацией путем бесконечных, устойчивых и необратимых рекомбинаций.
Таким образом, получив, как нам кажется, ответ на один из поставленных вопросов, и возвращаясь к началу данной главы, следует попытаться найти объяснения оставшимся: что есть парабиоз, физиологическая лабильность, доминанта, адаптационный синдром, функциональная система?
Прежде всего, каждый из этих терминов составляет суть вполне самостоятельных и фундаментальных теорий в физиологии и медицине. Поэтому в ответах на эти вопросы мы сочли необходимым в сжатой и, по возможности, в предельно чёткой форме попытаться изложить основные постулаты этих колоссов академической науки.
Основные постулаты учения профессора
Н.Е. Введенского (1852-1922) «О парабиозе и физиологической лабильности»:
В современном толковании «парабиоз»[3] это понятие имеющее двойное значение:
1. Реакция возбудимой ткани на воздействие раздражителей и характеризующаяся тем, что измененный участок нерва (мышцы) приобретает низкую лабильность и поэтому не способен к проведению заданного ритма раздражения;
2. Искусственное соединение двух (реже трех) животных через кровеносную систему или сращиванием их тканей для изучения взаимовлияний одного организма на другой.
В рамках предложенной темы изложения, нам предстоит более подробно рассмотреть понятие «парабиоз» в теории профессора Н.Е. Введенского, впервые представленного вниманию научной общественности в 1901 году.
По Н.Е. Введенскому: «Парабиоз, - это состояние, характеризующееся «стойким не колебательным возбуждением», представленное в развитии тремя стадиями: провизорной, парадоксальной и экзальтационной» (А.А. Ухтомский, 1950).
Цитируя профессора К.М. Быкова (1954) «парабиоз Введенского – это состояние углубившегося и утратившего способность распространяться возбуждения, представленное в развитии тремя стадиями: уравнительной, парадоксальной и тормозящей».
1. «Уравнительная» (по К.М. Быкову) или «провизорная» (по Н.Е. Введенскому) стадия – это исчезновение различий в действии слабых и сильных ритмических раздражений;
2. «Парадоксальная» стадия (по Н.Е. Введенскому и К.М. Быкову - на сильное раздражение возникает слабая реакция, в ответ на слабые раздражения – значительный ответ;
3. «Экзальтационная» (по Н.Е. Введенскому) или «тормозящая» (по К.М. Быкову) стадия – потеря способности отвечать на раздражение (рефрактерность или ретенция).
В связи с этими фактами, полученными Н.Е. Введенским в опытах на нервно-мышечном препарате, а позже замеченных им аналогий в деятельности центральной нервной системы, значение его работ не возможно переоценить. Они легли в основу:
1. электрофизиологических и биофизических эффектов (возбудимость, сократимость, ретенция рефрактерность, реституция, экстра- и постэкстрасистолическая потенциация);
2. нейрофизиологических эффектов (возбуждение, состояние покоя, состояние торможения, состояние запредельного торможения, состояние тотального и локального аффекта, состояние клеточного аффекта).
«Пессимум» по Н.Е. Введенскому, - это «деятельное успокаивание» (или иначе - порог возбудимости), а «оптимум» по тому же Н.Е. Введенскому - это норма возбудимости (двойной порог возбудимости).
«Фактор функциональной лабильности», о котором пишет Н.Е. Введенский - это «последнее максимальное число импульсов, которое ткань готова воспроизвести за секунду в точном соответствии с ритмом падающих на неё раздражений».
Фактически, опередив время на семьдесят с лишним лет, Н.Е. Введенский сформулировал суть срочного (немедленного) «функционального» и «энергетического» резерва любой «возбудимой среды». Параметра, введение которого в практику физиологов, клинических физиологов, научных работников сейчас, и который мы только пытаемся внедрить в жизнь сегодня, если он будет принят, способен качественно изменить будущность медицинской науки, биологии и естествознания.
В собрании сочинений А.А. Ухтомского (собр.соч.1966) можно найти: «…вывод, который следует из совокупности работ покойного[4] над возбудимыми элементами, но который он сам почему-то не пожелал сделать, а именно, что нормальное отправление органа (например, нервного центра) в организме есть не предопределённое, раз навсегда неизменное качество данного органа, но функция его состояния».
На наш взгляд, это прямое «введение» в концепцию профессора Ф.З. Меерсона о качестве «системного структурного следа», «долговременной и срочной фенотипической адаптации» (1981, 1988) и, одновременно, прямое указание на качество и состав собственно управляющего агента[5] этих естественных защитных (по Е.И Чазову 1996) или адаптационных (по H.Selye 1950) реакций организма[6].
«Введенский был глубоко убеждён, - пишет профессор А.А. Ухтомский (опубл. в 1950 году), что качественные различия результатов столкновения живого вещества с различными агентами могут и должны быть поняты, как производная от количества принципиально однородных зависимостей». Сегодня - это одно из основных положений современной патологической физиологии «адаптационно-приспособительных реакций в системе гомеостатической регуляции» организма (Н.А. Онищенко, 1975-2003).
Не боясь нарушить принцип краткости изложения, важно подчеркнуть то, что основные положения теории профессора Н.Е. Введенского, сформулированные автором, были изложены или его учениками, среди которых профессор А.А. Ухтомский[7], или его единомышленниками, каким был профессор К.М. Быков. Сам же профессор Н.Е. Введенский практически ничего не публиковал и, даже, не всегда и не всё, из полученного им материала, обрабатывал. Известно лишь очень небольшое количество работ, опубликованных самим автором. Нам же посчастливилось ознакомиться только с одной из них: «О соотношении между раздражением и возбуждением при тетанусе», опубликованной в Санкт-Петербурге ещё в 1886 году.
Очень приятно и торжественно, что сегодня имя профессора Николая Евгеньевича Введенского заслуженно поднято историей на пьедестал науки, действительно достойный его выдающихся заслуг.
Основные постулаты учения «О доминанте» профессора А. А. Ухтомского (1875-1942):
«Доминанта» есть естественное
и неотъемлемое детище школы Введенского
А.А. Ухтомский
В основу учения «О доминанте» был положен главный принцип общего правила работы нервных центров, который А.А. Ухтомский назвал «Принципом доминанты».
В авторском изложении, он формулируется следующим образом: «Возбуждение, рождающееся от раздражения, имеет тенденцию направляться к центру наиболее деятельному». Это положение профессора А.А. Ухтомского было противопоставлено двум основным концепциям «общего правила» и завоевало в то время свой приоритет и признание. Это:
1. принцип E. Hering-Breuer’a: «Возбуждение, рождающееся от раздражения, имеет тенденцию разливаться по нервным центрам так, что имеющаяся в данный момент реакция, переводится в противоположную» и
2. принцип Uexkill’я: «Возбуждение, рождающееся от раздражения, имеет тенденцию направляться всегда к центру наиболее покоящемуся».
«…доминанта есть не привилегия высших нервных этажей, но общий принцип нервных центров», - пишет в ответ на это профессор А.А. Ухтомский (1924).
«…принцип доминанты…- общий modus operandi центральной нервной системы» (собр. соч. 1966), а собственно «…доминанта характеризуется своею интуицией…, однажды вызванная, она может восстанавливаться» (1924) (прямое указание на «системный структурный след» в теории профессора Ф.З. Меерсона).
В то время в физиологии ещё не существовало таких понятий как: «специфический» физиологический ответ, «неспецифическая» или «типическая» физиологическая реакция. Но уже тогда профессор А.А. Ухтомский, пытаясь объяснить механизмы «адаптивно-приспособительных реакций», о которых также пока ничего не было известно, ввел не закрепившееся сегодня понятие «физиологический орган», вложив в него следующий смысл: «это, прежде всего, механизм с определенным однозначным действием». Он писал: «Громадное многообразие морфологических черт, которые мы открываем в том или ином образовании нашего тела, впервые приобретает значение «физиологического органа»… Именно физиологическая равнодействующая даёт комплексу тканей значение органа как механизма».
По существу, это блестящее предвидение, нашедшее себя в дне сегодняшнем. Далее он говорит: «Однажды начавши усиленно работать, нервная система на высоте своего действия вовлекает в сферу работы организма всё новые и новые порции энергии со стороны. Лишь как о вторичном, постепенном достижении можно говорить о прогрессирующей экономике каждой отдельной центральной функции, но она не есть нечто, данное с самого начала, как роковое последствие наименьшего действия. Это достижение, дающееся, может быть, многими годами. Может быть иногда недостижимое в течение целой жизни». (Из стенограммы выступления на заседании биологического студенческого научного кружка Ленинградского университета от 2.04.1927).
Таким образом, формулируются основные черты и свойства «доминанты» в теории профессора А.А. Ухтомского (1926-1927):
1. повышенная возбудимость;
2. стойкость возбуждения;
3. способность к суммированию возбуждений;
4. инерция – способность удерживать и продолжать в себе ряд начавшегося возбуждения и тогда, когда первоначальный раздражитель уже миновал;
5. «Пережитая доминанта» при затухании не аннулируется, а тормозится во времени. Воспитывать данную доминанту, тщательным образом обихаживать её, следить за тем, чтобы она не перевозбудилась, не перешагнула известной величины…»
Для нас остаётся и, по всей видимости, останется загадкой, каким образом столь блистательная по своей сути теория «О доминанте» не смогла учесть уже известных в то время позиций «Учения о возбуждении и торможении» ещё одного выдающегося русского физиолога профессора Ивана Михайловича Сеченова.
Возможно, тому виной изначально кажущаяся «несводимость» описываемых процессов. Хотя очевидно, что для такого уровня учёных, какими были профессора Н.Е. Введенский и А.А. Ухтомский, вопрос об установлении однотипности реакций со стороны возбудимых тканей при их раздражении, на наш взгляд, не мог бы составить больших трудностей.
Возможно, каким-то образом повлиял официальный арест книги И.М. Сеченова в связи с её «неоспоримо вредным направлением[8]», слишком смело проникающим в темную до того времени область психических явлений сознательного и бессознательного, позже возведённых профессором Зигмундом Фрейдом на пьедестал психофизиологии.
То ли сложный характер и замкнутость профессора Н.Е. Введенского, передавшиеся его ученику, то ли разобщенность научных и профессиональных школ сыграли определенную роль, но вышедшая в свет в 1863 году сначала в журнале «Медицинский вестник» (в сокращенном варианте), а затем в 1866 году отдельным изданием известная всему миру работа И.М. Сеченова «Рефлексы головного мозга», раскрывающая суть учения о рефлекторных механизмах не только возбуждения, но и торможения, не была упомянута в своих трудах ни профессором Н.Е. Введенским, ни профессором А.А. Ухтомским.
Суть же данного пояснения в том, что будь принципы, заложенные профессором И.М. Сеченовым в основу физиологии «возбудимых сред», приняты «Учением о доминанте», работа немецкого биолога H. Dotterweich «О привыкании к раздражению, адаптации и торможению», опубликованная в 1940 году, не вызвала бы столь мучительных переживаний со стороны профессора А.А. Ухтомского.
Суть ремарки «О привыкании к раздражению, адаптации и торможению» немецкого биолога H. Dotterweich (1940), принципы возбуждения и торможения в учении профессора И.М. Сеченова (1829-1905)
«Мы приобрели для могучей власти
физиологического исследования вместо половинчатого
весь нераздельно животный организм»
И.П. Павлов
Из собрания сочинений А.А.Ухтомского (1962, т.6, с.168) мы узнаем, что профессор очень болезненно, но как истинный ученый, более чем достойно, прореагировал на настоящую ремарку H. Dotterweich (1940). Он с горечью и, вместе с тем, каким-то воодушевлением, писал: «Некоторые признаки побуждают думать, что современная нам физиология и биология вообще начинают переживать потребность в критическом пересмотре своих ведущих понятий и рабочего идейного инвентаря. Это – знамение времени…. Сейчас я имею ввиду поучительную книгу дрезденского биолога Х. Доттервейха, посвященную пересмотру концепции равновесия в современных биологических науках».
В чём же существо этой ремарки, заставившей учёного с мировым именем, «пересмотреть» некоторые свои признанные научные убеждения, которые до того оставались неколебимыми даже работами профессора И.М. Сеченова?
H. Dotterweich пишет буквально следующее: «Очень краткие раздражения при частом повторении не вызывают более реакции. Это обыденное наблюдение и процесс, при посредстве которого животное защищается от излишних влияний среды». Это явление автор назвал – «адаптацией и торможением вместе, или иначе, - привыканием к раздражению» (1940).
Комментируя работу H. Dotterweich, профессор А.А. Ухтомский писал: «Адаптация понимается автором как приспособление к монотонному изменению среды. «Адаптация», говоря вообще, есть привыкание или приспособление (Angleichung oder Anpassung) к новому непрерывно продолжающемуся состоянию раздражения, т.е. к некоторым более длительно продолжающимся изменениям определённых условий среды. Отсюда следует, что процесс адаптации есть не что иное, как процесс равновесный, при котором определенные компоненты организма (например, рецептивные клетки и органы чувств) изменяются в направлении нового равновесия, отвечающего изменённому условию среды».
Огорчение профессора глубоко понятно, хотя общие идеи ничуть не были опровергнуты в главных своих постулатах. Да, не накопление раздражения, а привыкание к ним, тренинг, адаптация. Но что более удивительно, ведь слова, произнесенные А.А. Ухтомским «в комментариях» по этому поводу, - это фактически, прямое указание на готовность организма, тканей к изменениям, на способность его сохранить баланс в обмене веществом, энергией и информацией с внешней средой в процессе адаптации. А это, - материалы современных работ наших коллег и нас в 1995-2004 годах.
Далее профессор А.А. Ухтомский продолжает: «При очень медленных перестановках равновесия и самые инерционные элементы организма претерпевают сдвиги, ибо принципиально они не отличаются от прочих компонентов. Требуется лишь адекватность внешнего, наружного и внутреннего процессов во времени протекания. Изменяться должны и вещества ядер в клетках и гены».
Позиция, нашедшая яркое подтверждение через 45 лет в работах профессора Ф.З. Меерсона, посвященных «запоминанию», «тренингу» организма к факторам стресса из внешней среды на генетическом уровне, получившая название «системного структурного следа».
«Адаптивное значение мутации по отношению к внешним для организма условиям является в известной степени даже невероятным, потому что специфические изменения генов производятся не непосредственно внешними раздражителями среды, но в первую очередь, физиологическими раздражителями, которые возникают вторично по поводу внешних влияний». «Мутабильность» – «классическая инерция»; нарушений равновесия уже нет, а мутация продолжается» (А.А. Ухтомский).
Остаётся только недоумевать и восхищаться возможностями «не инструментального», а интуитивного понимания сути физиологических процессов. Тем, что сегодня мы называем возможностью научного предвидения. Ведь как точно обозначен ключевой момент в тончайших механизмах адаптации, названный в 1975 году «фактором энергетической задолженности и энергетического дефицита» (В.И. Шумаков, Е.Ш. Штенгольд, Н.А. Онищенко 1975), который корректнее, наверное, следовало бы назвать «фактором энергетического дисбаланса». Но об этом позже.
Основные положения «Учения об адаптационном синдроме» профессора Hans Hugo Bruno Selye (1950)
«Адаптационный синдром сам по себе
не является патологической реакцией;
наоборот, это необходимая физиологическая реакция
на повреждение как таковое, имеющая защитный характер»
Ганс Селье
В своей книге «Стресс без дистресса» (1979) профессор Ганс Селье пишет: «Почти четыре десятилетия я изучал в лаборатории физиологические механизмы приспособления к стрессу и убедился, что принципы защиты на уровне клетки в основном приложимы и к человеку и даже к целым сообществам людей».
В другой своей известной книге «Очерки об адаптационном синдроме» (1960) профессор сам формулирует основной «лейтмотив» своего учения «об адаптационном синдроме». Он пишет: «Ключ к пониманию и оценке концепции адаптационного синдрома нужно искать не в области фактов, а в области представлений, на которых она основана. Следует постоянно помнить, что эта концепция покоится на нескольких простых положениях, иллюстрируемых прилагаемой схемой (мы её приводим также «дословно» на рисунке 1).
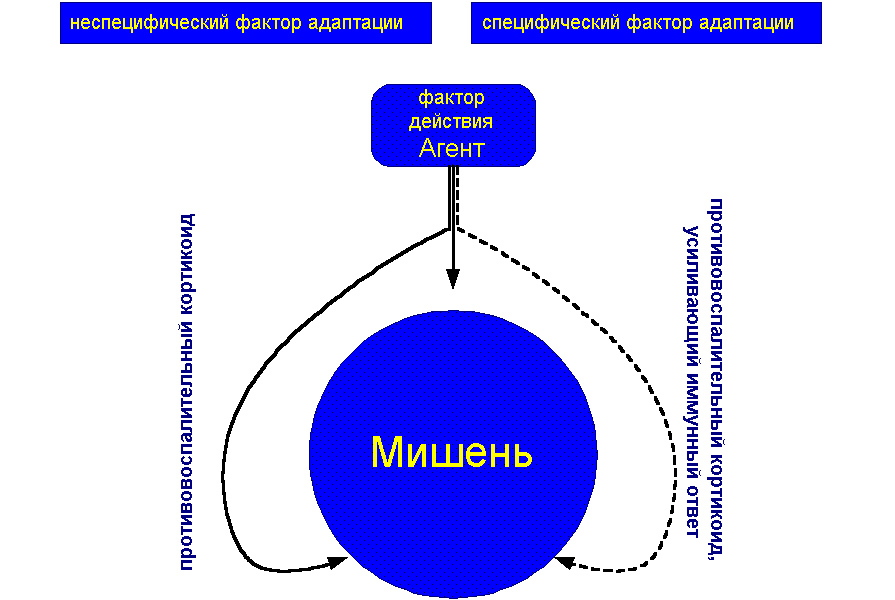
Рисунок 1. Основные положения, иллюстрирующие концепцию адаптационного синдрома по Hans Hugo Bruno Selye (1950)
Это поможет вам уследить за моим сложным рассказом о стрессе. Все агенты, которые действуют на организм или какую-либо часть его («мишень») вызывают двоякого рода эффекты:
1. специфические (пунктирная стрелка), например, эфир вызывает анестезию, тифозная салмонелла – паратиф;
2. неспецифические стереотипные эффекты стресса, которые проявляются в виде общего адаптационного синдрома (трёх ветвистая стрелка). Стресс прямо воздействует на «мишень»; это может вызвать местные тканевые повреждения, например ожог или прямое влияние микробов на клетки (прямая ветвь толстой стрелки), но некоторые его эффекты опосредуются через гуморальные или нервные пути (изогнутые ветви толстой стрелки).
Главный гуморальный путь идет через переднюю долю гипофиза и корковый слой надпочечников, сущность участия которых в этих реакциях заключается в секреции ими «адаптивных» гормонов. Среди них, с одной стороны – СТГ и противовоспалительные кортикоиды (например, ДКА), соответственно подавляющие и стимулирующие воспалительные реакции на повреждение.
Основным видом реакции организма на местный стресс является воспаление, на системный стресс – шок. Различные комбинации этих двух видов реакций составляют сущность большинства болезней.
Тканевая реактивность, регулируемая к примеру адаптационными гормонами, часто определяет, поддаётся ли организм болезни или устоит против воздействия потенциально патогенного агента, используя свою способность к адаптации. В синдроме адаптации участвуют и другие гуморальные агенты, а также нервная система, однако природа и степень их участия ещё недостаточно выяснены, чтобы их можно было детально рассмотреть в настоящее время». Конец цитаты.
Уже в этой главе мы постараемся восполнить этот «пробел», о котором пишет профессор Г. Селье, основными постулатами «общей теории функциональных систем» академика П.К. Анохина (1958-1996) и некоторыми положениями концепции профессора Ф.З. Меерсона «о системном структурном следе в общем адаптационном синдроме» (1965-1988). Это, пожалуй, один из немногих «пробелов» в учении профессора Г.Селье, замеченный ещё академиком М.Г. Дурмишьяном в 1960 году: «В значительной мере это объясняется тем, что проблема в целом изучалась без должного учёта роли нервных механизмов», - пишет академик в своей статье «Стресс и нервизм» (М: Медгиз, 1960, стр. 17).
По нашему твердому убеждению, - это не недостаток теории профессора Г.Селье, поскольку никак не умоляет её основополагающих практического и теоретического достоинств. Но «насильно мил не будешь», а «объять необъятное» не только трудно, часто – нельзя «да и надобности нет».
По всей видимости, синдром «болезни» не «объять необъятное» был всегда и, что несколько печально, в нашей же области науки, и в той же связи. О нём было упомянуто чуть выше в этой же главе при рассмотрении «Учения о доминанте» профессора А.А. Ухтомского, оставившего без внимания механизмы, раскрытые «Учением о возбуждении и торможении» профессора И.М. Сеченова для центральной нервной системы и со ссылкой на поправку доктора H. Dotterweich, «поколебавшую» устои физиологии в области адаптации.
Суммируя основные выводы своей теории, профессор Г. Селье пишет: «Все эффекты биологических агентов и в особенности гормонов во многом обусловливаются «кондициональными факторами» (затушеванная окружность на рисунке 1); одни из них являются экзогенными (диета, температура, освещение), другие – эндогенными (наследственность, конституция, предыдущие воздействия). Таким образом, - обобщает автор, - состояние стресса реализуется в организме посредством многочисленных механизмов, его проявление всегда маскируется разнообразными специфическими эффектами воздействующего агента и «кондициональными факторами», т.е. «почвой», которая различна в каждом индивидуальном случае. Такие обусловливающие факторы могут избирательно затрагивать те или иные механизмы развития стресса. Этим и объясняется то, что единое, в сущности, состояние стресса меняет своё проявление в различных случаях.
Адаптационный синдром сам по себе не является патологической реакцией; наоборот, это необходимая физиологическая реакция на повреждение как таковое, имеющая защитный характер. Однако этот синдром, подобно любой другой биологической реакции, не всегда оказывается оптимально эффективным. Несовершенство его (например, абсолютный или относительный избыток или недостаток одного или нескольких адаптивных гормонов) играет важную роль в патогенезе большинства болезней. Заболевания, при которых неадекватность синдрома адаптации имеет даже большее значение, чем специфические эффекты патогенного агента, относятся преимущественно к «болезням адаптации». Конец цитаты.
Еще раз хочется подчеркнуть, что в любом случае: «за» это или «против», но изначальная мысль автора всегда лучше её интерпретации. Тем более, что написать лучше, яснее и доступнее, чем директор монреальского Института экспериментальной хирургии и медицины, профессор Ганс Селье, если не нельзя, то очень трудно, и, во всяком случае, - «да и надобности нет».
Основные положения концепции профессора В.В. Парина и теории профессора
Ф.З. Меерсона «О системном структурном следе в общем адаптационном синдроме» (1965-1988)
Как-то в одной из своих университетских лекций профессор Ф.З. Меерсон сказал: «По существу, вопрос о механизме фенотипической адаптации состоит в том, каким образом потенциальные генетически детерминированные возможности организма в ответ на требования среды преобразуются в его реальные возможности».
Это, действительно, ключевой вопрос, ибо без знания механизмов, поддерживающих гомеостаз в условиях постоянно меняющейся внешней среды, любая теория, даже самая совершенная, носит лишь описательный характер и вряд ли применима на практике.
Профессор Феликс Залманович Меерсон один из первых учёных, кто всерьез обратился к системному и последовательному изучению механизмов гомеостатических отношений на уровне организм-клетка, клетка-организм, применив для этого весь доступный ему арсенал методов исследований.
Разрабатывая «Теорию индивидуальной адаптации к среде» он выделяет следующие основные её положения:
1. «В развитии большинства адаптационных реакций определённо прослеживаются два этапа: начальный этап срочной, но несовершенной адаптации и последующий этап совершенной долговременной адаптации.
2. Срочный этап адаптационной реакции возникает после начала действия раздражителя и, следовательно, может реализоваться лишь на основе готовых, ранее сформировавшихся физиологических механизмов.
3. Наличие готовой функциональной системы или её новообразование само ещё не означает устойчивой, эффективной адаптации.
4. Для перехода срочной адаптации в гарантированную долговременную, внутри возникшей функциональной системы должен реализоваться некоторый жизненно важный процесс, обеспечивающий фиксацию сложившихся адаптационных систем и увеличение их мощности до уровня, диктуемого средой.
5. Адаптация организма к различным факторам среды формирует системные структурные следы различной архитектуры.
6. Основой относительной специфичности фенотипической адаптации является тот факт, что разветвленный системный структурный след, составляющий основу адаптации к определенному фактору, нередко имеет в своем составе компоненты, которые могут повышать резистентность организма к действию других факторов». Конец цитаты.
7. Стадийность индивидуальной «адаптации - деадаптации»:
· 1-ая стадия «срочной адаптации»;
· 2-ая стадия «перехода срочной адаптации в долговременную»;
· 3-я стадия «сформировавшейся долговременной адаптации»;
· 4-ая стадия «истощение».
Схематически основные механизмы индивидуальной адаптации представлены на рисунках 2 и 3, а её стадии на рисунках 3 и 4 в полном соответствии с теорией профессора Ф.З. Меерсона и заимствованы из его работ.
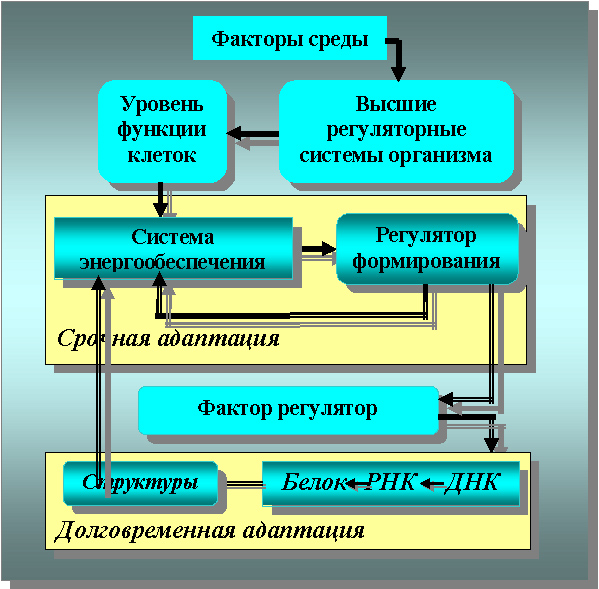
Рисунок 2. Схема клеточного звена долговременной адаптации
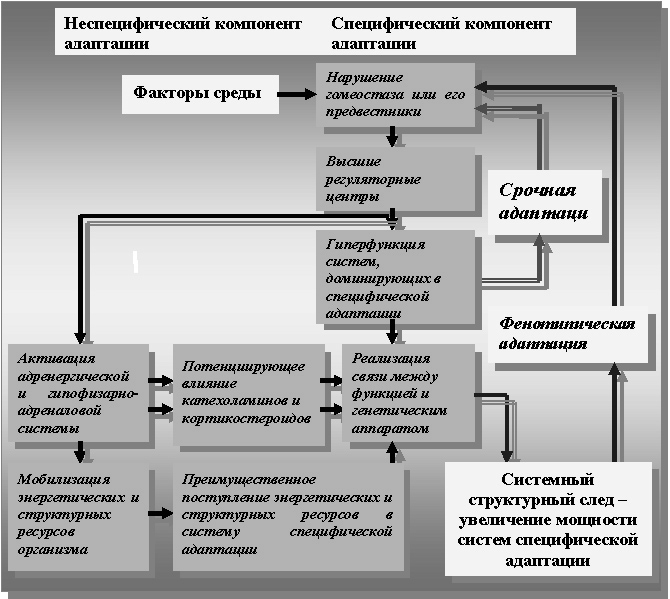
Рисунок 3. Основной механизм и стадии фенотипической адаптации
Принцип профессора Н.А. Онищенко: «Энергетическая задолженность и энергетический дефицит» в общем генезе адаптационно-приспособительных гомеостатических реакций
(1975-1988)
«За любой эффект устойчивости надо платить…»
Н.А.Онищенко

Если профессор Г. Селье обнаружил и блестяще описал «Стресс-синдром», а профессор Ф.З. Меерсон получил ясное и убедительное понимание того, «что из этого произойдет» и «как это будет происходить», то профессор Нина Андреевна Онищенко установила и последовательно доказала этиологию и патогенез происходящего.
Вообще такие понятия как «энергетическая задолженность» и «энергетический дефицит» были предложены профессором Н.А. Онищенко ещё в 1970-1972 году. Но более или менее весомо они впервые прозвучали в 1975 году, попав в лексикон трансплантологов, а затем физиологов, клинических физиологов и патофизиологов. Это произошло вместе с выходом в свет книги профессора М.Е. Райскиной, Н.А. Онищенко, и Б.М. Шаргородского с соавторами «Методы прижизненного исследования метаболизма сердца»[9] и монографии профессора В.И. Шумакова, Е.Ш. Штенгольда и Н.А. Онищенко[10] «Консервация органов».
В этих работах впервые описание процессов, происходящих в «умирающем органе» (сердце, почка), ожидающем трансплантации, сопровождалось физически выдержанной и математически четкой калькуляцией реального энергетического баланса и энергетических потерь.
Трудно сказать, что названные понятия прочно вошли в обиход коллег (дискуссия все ещё продолжается), но их значение во истину неоценимо. В замен привычному, но увы, не вполне чёткому понятию «потребление кислорода» и «кислородный баланс», выражаемых в объемных единицах, появились физически ясные и грамотные «неожиданности и непривычности» для представителей медицинских и биологических наук: «энергетическое обеспечение», «энергетическая задолженность», выраженные в калориях, отнесённых к граммам массы, «скорость накопления энергетического долга» в кал/мин.
Оппоненты по сей день, весьма часто возражают против использования этих терминов, и во многом с ними нельзя не согласиться, т.к. «энергетическая задолженность» – это величина «расчётная». Но, как мы имели честь и возможность показать, эволюционный процесс необратим всегда и везде, в том числе, и в науке. Профессор Н.А. Онищенко катализировала необратимость «рекомбинации» консервативного мышления своих коллег, будучи по существу абсолютно правой.
Всё дело и вся сложность в том, что в генезе повреждения, стресса или любого другого действия на гомеостатическую систему, о которых мы намерены говорить ниже, «кислородный долг» и «дефицит кислорода» имеют место быть, но они не составляют обобщенную суть большинства физиологических и патофизиологических механизмов. «Дефицит кислорода», если в качестве примера говорить о нём, это только «повод» возникновения цепи последующих гомеостатических реакций. Повод и причина появления дефицита энергии. Бесспорно, дефицит кислорода самостоятельно вызывает энергетический дефицит. Но это ни в коем случае не означает, что энергетический дефицит является следствием только недостатка кислорода. Провоцирующих причин, следствием которых является недостаток в энергетическом обеспечении адаптивно- приспособленных состояний гомеостатических систем (в частности – живых клеток, организма) можно назвать очень много. Например: травма, инородные включения, трансплантаты, бактериальное заражение, токсикоз, гиперфункционирование, гипо- и гипертермия и т.д.
Понятие «энергетический дефицит» или «дефицит энергии» принципиально решили главную проблему: понимание генеза адаптационно-приспособительных реакций. Иллюстрируя сущность принципа «энергетического дефицита» в общем генезе адаптивно-приспособительных гомеостатических реакций, досконально исследованных профессором Н.А. Онищенко и её сотрудниками, следует обратиться к схеме одного из её учеников и последователей профессора В.И. Кирпатовского (рисунок 5).
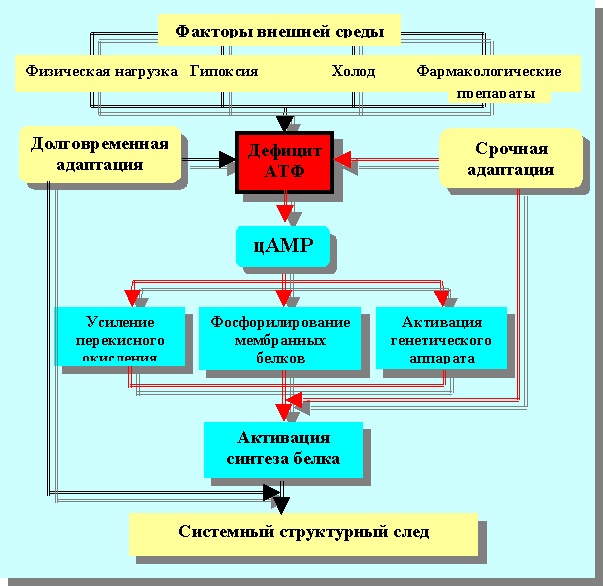
Рисунок 5. Схема клеточного звена фенотипической адаптации по В.И. Кирпатовскому
Хочется особенно подчеркнуть, что колоссальная роль и вклад в развитие гомеостатического учения работ профессора Н.А. Онищенко совершенно бесспорны в том, что ей удалось выделить главный и всеобщий компонент собственно типической или неспецифической физиологической реакции, дифференцировать причинно следственную совокупность сложнейшего по сути действия, каким является адаптационный синдром[11].
Пример 1. На уровне гомеостаза клетки - это нарастающее несоответствие (дефицит) скоростей производимой и потребляемой энергии ишемическим миокардом, отслеженный и общей кинетикой теплопотерь, изменениями абсолютных [ATP], [АDP], [Pi], в особенности, - относительных величин макроэргических соединений внутри клеток (т.е. их соотношений: [ATP]·[АDP]·[Pi]-1) и динамикой уровня поляризации мембран, механического напряжения покоя (тонуса) миокарда, как допустимых критериев в оценке жизнеспособности искусственно остановленного сердца. Именно посредством первичного применения расчётов условного показателя «дефицит энергии», как разницы между должными и истинными энерготратами, удалось, в конечном итоге, уйти от этого абстрактного показателя в область традиционных величин, соотносящих скорость производимой и потребляемой энергии через показатели эффективности функционирования системы[12].
Пример 2. На уровне органа был установлен абсолютно идентичный диагноз, также построенный на принципах расчёта энергетического дефицита и эффективности функционирования при его искусственной изоляции. Он позволил выявить не только банальный генез умирания работающего и несущего функциональные нагрузки сердца, развивающегося в результате одной только механической изоляции, что серьёзно разрушило представление о «перспективах» длительного сохранения сердца вне организма в работающем состоянии и «физиологичности» такого рода методов. Было убедительно показано, что при внешней видимости и общепринятой объективной констатации удовлетворительного состояния сердца, включая стабильность его гемодинамических и электрофизиологических показателей вплодь до 11-12 часов работы вне организма, уже в первые 60-120 минут после его изоляции, развивается настолько мощный «дефицит» энергии, что миокард не справляется с ним даже путём перераспределения статей своего энергорасхода. Один-два часа такого напряжения в энергетическом метаболизме, когда эффективность функционирования структур миокарда свидетельствует о колоссальной гиперфункции, наступают настолько необратимые изменения, что они наверняка гарантируют гибель сердца, хотя и по-разному пролонгированную, но наступающую при любых обстоятельствах. Подавляющее же большинство коллег, занимавшихся изучением этого способа консервации органов, до появления таких сообщений было твёрдо убеждено в обратном8.
Пример 3. На уровне системы органов, а таковой была избрана весьма популярная в 1950-1970 годах система изолированного сердечно-лёгочного комплекса, как наиболее «физиологичная» с точки зрения гемодинамики и методики забора для искомых «условий оптимизации» способов консервации сердца. Но и на этой модели была установлена абсолютно идентичная кинетика умирания, достоверно повторяющая и динамику и сроки исследованных ранее нарушений, связанных только с несоответствием скоростей производимой и потребляемой энергии и, конечно, через показатели эффективности функционирования самой системы8.
Пример 4. По существу, опираясь на «принцип Н.А.Онищенко», сотрудникам её лаборатории удалось очень подробно расшифровать весь механизм действия и самих консервирующих растворов, применяемых в трансплантологии и кардиохирургии. Это, в свою очередь, указало главное, ключевое звено повреждения сохраняемого органа уже на самом раннем этапе манипуляций с ним. На стадии отмывания сосудистого русла от крови консервирующим раствором. В особенности, это подробно было исследовано на миокарде. В результате, была разработана некая стратегия защиты, контролируемая по эффективности исполнения. В частности, только путём контролируемого устранения неблагоприятных последствий от введения кардиоплегических растворов в коронарное русло, а это всегда сопряжено с повышением тонуса сердца (калиевая контрактура или контрактура деполяризации), а значит, лишним расходом энергии, удалось достичь эффективной защиты миокарда на время всего хирургического вмешательства, без использования дополнительных перфузий, отвлекающих хирурга и персонал, не говоря уже о сокращении времени самого вмешательства. А контроль за эффективностью такого действия позволяет сделать режим отмывания коронарного русла индивидуальным[13].
Пример 5. Более того, зная и контролируя генез дополнительного повреждения, вызванного самим хирургическим вмешательством и процедурами, сопровождающими его, открыт грандиозный плацдарм для получения наиболее благоприятных результатов, подтверждённых уже десятилетней статистикой. Только опираясь на «принцип Н.А. Онищенко», удалось в полной мере разработать и сформулировать общий подход к выбору индивидуальных средств и способов избираемой стратегии защиты миокарда оперируемого сердца, а также организма пациента во время операции и в дооперационный период[14].
Пример 6. Время и настойчивость показали, что «принцип Н.А. Онищенко» применим и равно корректен на уровне гомеостаза всего организма. Так при расчёте энергетических задолженностей и эффективности функционирования всей кровеносной системы ad mass во время исполнения искусственного кровообращения в связи с коррекцией приобретённой патологии сердца, удалось вполне определённо установить некое скрытое несоответствие в традиционной тактике перфузии потребностям организма[15] пациента. Это, в свою очередь позволило найти подход и выработать алгоритм для адаптации режима экстракорпоральной перфузии аппарата искусственного кровообращения к индивидуальным особенностям кровеносной системы каждого конкретного пациента[16].
Яркой иллюстрацией принципиальной важности и правильности действия «принципа Н.А. Онищенко» на двух важнейших уровнях гомеостатического устройства (клетка, организм) может служить его универсальный прикладной эффект. Он нашёл себя при защите сердца от ишемии и гипотермии во время реконструктивных операций на клапанах и коронарных сосудах сердца. Десятки кардиохирургических клиник Москвы и Санкт-Петербурга (профессор В.И. Шумаков и его сотрудники (1978-1996), профессор М.Л. Семеновский и сотрудники (1985-1998), профессор А.Б. Зорин и сотрудники (1985-1990), профессор В.К. Новиков и сотрудники (1988-2003) и многие другие) используют методологию кардиоплегии, составленную с учетом требований теоретической базы школы профессора Н.А. Онищенко, к коей себя имеет честь относить и автор этих строк.
Трансплантация почек, консервация сосудов, печени, гепатоцитов, селезенки, спленоцитов, b-панкреацитов и многих других жизненно важных органов, тканей, клеток (В.И. Шумаков и соавт. 1977-1999, И.Д. Кирпатовский и соавт. 1980-2003, Н.А. Лопаткин и соавт. 1989-2003) производится сегодня по методикам, разработанным сотрудниками лаборатории консервации органов, возглавляемой профессором Ниной Андреевной Онищенко.
Ещё более замечательным свойством «принципа Н.А. Онищенко», являющимся предметом изучения и теперь, можно назвать его распространимость не только на объекты, представительствующие клеточный и организменный уровень гомеостаза. Как это ни удивительно, родившись в недрах чисто прикладного использования медицинского назначения, он может быть блестяще применим на элементарном уровне организации материи.
Пример 7. Так, если рассмотреть численный ряд всех известных сегодня науке элементов периодической системы Д.И. Менделеева, разместив их в полном соответствии принципу, избранному ещё самим Дмитрием Ивановичем, т.е. по возрастанию масс, то будет получен некий график хорошо известной прямолинейной зависимости атомных весов и атомных номеров элементов в данном воспроизведённом ряду (рисунок 6, верхняя область диаграммы). Если же применить к нему (этому ряду) «принцип Н.А.Онищенко» и рассчитать суммарное количество так называемых «вакантных» электронных мест[17] для каждого атома элемента, то получим новый ряд чисел, автоматически распределяющий весь известный элементный состав атомов веществ в известные семь периодов (рисунок 6). Более того, вновь образованный численный ряд полностью отвечает неким известным характеристикам свойств элементов, изменяющихся внутри каждого периода. А ведь по сути, если трактовать этот эффект с авторских позиций «принципа Н.А. Онищенко» - это есть ни что иное, как некий абстрактный «электронный дефицит». На языке же химии – это «электронная вакансия».
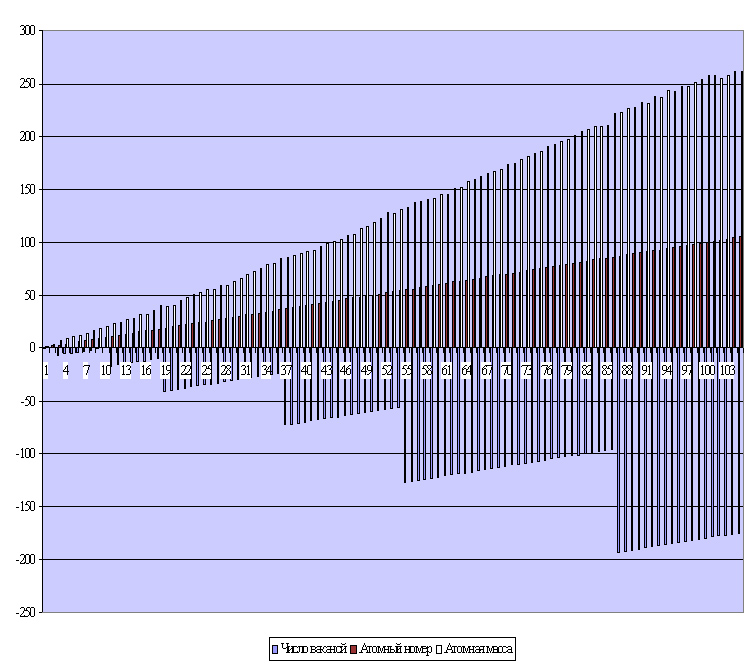
Рисунок 6. Число электронных вакансий (нижняя область диаграммы) в периодической системе элементов Д.И. Менделеева в соответствии с принципом Н.А. Онищенко
В заключение этого важного для изложения раздела, хочется привести цитату самой профессора Н.А. Онищенко, посвятившей всю свою научную жизнь решению задач трансплантологии и противоишемической защиты органов. Говоря о способах коррекции ряда физиологических состояний сердца и почек, она пишет: «Фармакологическая адаптация выражается в первоначальном усилении клеточных энергозатрат, благодаря чему повышается резистентность структур к повреждению» (1996), т.е. «за любой эффект устойчивости, как и вообще за всё, надо платить». Конец цитаты.
Принцип профессора В.А. Фролова, дифференцированный подход к состоянию гиперфункции в «Теории патологии сердца» (1975-1988):
Применив принцип профессора Н.А.Онищенко при оценке «энергетической задолженности» миокарда через условный коэффициент K[18], характеризующий «результирующую синтеза и потребления энергии» кардиомиоцитами, профессор В.А.Фролов (1982) продемонстрировал блестящую систематизирующую возможность академического подхода к описанию патогенетических процессов, указав «принципиально возможные пути развития патологических процессов в сердце» (рисунок 7).
В своей монографии, подготовленной совместно с профессором В.С.Пауковым, профессор В.А. Фролов, комментируя приведенную на рисунке 6 схему, пишет: «В стадии инициальной гиперфункции сократительного миокарда патогенный фактор вызывает его относительную или абсолютную перегрузку, ведущую к усилению механической функции сердечной мышцы, что сопровождается повышением её потребности в энергии. Так как энергообеспечение ещё отстает от потребностей миокарда, появляется энергетический дефицит и включаются механизмы стимуляции энергообразования.
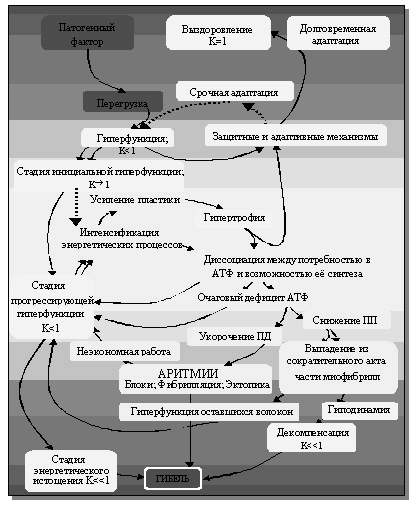
Рисунок 7. Схема развития патологических процессов в сердце по В.А.Фролову
P.S. Небольшие, но показавшиеся логически уместными, добавления внесены в схему пунктирной линией
Возникает гиперфункция митохондрий, выработка энергии повышается и, если в этот период произойдет устранение перегрузки кардиальных структур, возможно выздоровление». Конец цитаты.
По всей видимости, лучше проиллюстрировать процесс «срочной адаптации» языком физиолога, чем это сделал профессор В.А. Фролов, невозможно. Появившееся же понятие «инициальная гиперфункция» более чем убедительно демонстрирует и этап и характер этапа собственно генеза гиперфункционирования системы.
Далее профессор В.А. Фролов пишет: «Если же перегрузка ультраструктур мышечных клеток сердца не устранена, развивается вторая стадия – прогрессирующей гиперфункции сократительного миокарда и дальнейшее течение заболевания зависит от интенсивности перегрузки, а также от скорости и объёма включения снижающих её компенсаторно-приспособительных механизмов. В этой стадии патологические процессы развиваются по принципу порочного круга. При чрезвычайных перегрузках почти вся энергия расходуется на обеспечение механической функции, а ресинтетические процессы энергией не обеспечиваются. Наступает быстрое истощение ультраструктурных элементов и их гибель. Если степень перегрузки позволяет включиться в процесс компенсаторно-приспособительным механизмам, обеспечивающим и высокий уровень биосинтеза, заболевание пролонгируется…. В заключительной третьей стадии заболевания… уровень энергетического обеспечения миокарда уже не может вернуться к исходному значению. Выздоровление на этой стадии не возможно». Конец цитаты.
Основные положения «Структурных основ адаптации и компенсации нарушенных функций» профессора Д.С. Саркисова (1987-1988)
«Осмелится ли кто-нибудь утверждать,
что нужно различать законы жизни
в патологическом состоянии
от законов жизни в нормальном?»
Клод Бернар
«Первая общая черта компенсаторно-приспособительных реакций организма состоит в том, что все они представляют собой ту или иную комбинацию его физиологических функций;
Вторая черта, объединяющая компенсаторно-приспособительные реакции организма… заключается в том, что обычно они характеризуются усилением функций, из которых комбинируется каждый из них;
Третья общая черта, свойственная всем компенсаторно-приспособительным реакциям, заключается в том, что все они развёртываются на принципиально единой, стереотипной материальной основе; Таким образом…, адаптация – это, прежде всего изменение скоростей биологических реакций. Компенсаторно-приспособительные реакции, обеспечивающие гомеостаз, не являются какими-то особыми реакциями организма, а представляют собой разнообразные комбинации его физиологических функций, развёртывающихся на той же, что и в норме, материальной основе и, как правило, с большей, чем обычно, интенсивностью». Конец цитаты.
Вопрос о надежности биологических систем: «Современные исследования показывают, что это обеспечение (диапазон приспособительных колебаний интенсивности физиологических функций организма) строится на нескольких основных принципах:
1. В состоянии относительного функционального покоя из общего числа одноименных структур активно функционирует лишь какая-то часть.
2. При более или менее длительной функциональной нагрузке, когда оказывается недостаточным включение в активную работу даже всех структур, которыми располагает орган, происходит увеличение их числа, т.е. гиперплазия, (названная профессором Д.С.Саркисовым: «компенсаторная гиперплазия») количественно соответствующая уровню возросшей нагрузки.
3. Приспособительные колебания функциональной активности органов материально обеспечиваются не просто изменениями числа функционирующих (интенсивно обновляющихся) структур, но изменениями, происходящими в строгом временном соответствии между частотой и силой воздействия раздражителя». Конец цитаты.
Особенности гиперпластических процессов:
1. «Экстренная мера – усиление синтеза РНК; радикальная, долгосрочная мера – усиление синтеза ДНК.
2. Какой бы ни была форма адаптации в каждом конкретном случае, будь это даже самая срочная, практически немедленная адаптация, она никогда не осуществляется на чисто функциональной основе, всегда имея под собой соответствующую материальную базу».
3. ...всегда имеется гиперплазия не индифферентных структур, а строго специфически ориентированных в своём действии на нейтрализацию… того патогенного фактора, который индуцировал гиперплазию.
4. «Полифункциональность клеток» – ещё один механизм компенсации дефицита функции за счёт гетерогенности последовательно «отдыхающих» клеток и структур.
5. Способность организма к экономии материальных ресурсов и максимальной концентрации их на главном участке развёртывания приспособительных реакций – принцип перераспределения нагрузок и функций, как залог резервных возможностей компенсаторно-приспособительных реакций.
6. Принцип «антагонистической регуляции функций» – двойной контроль ослабляющих и усиливающих систем, соотношение которых определяет уровень функциональной активности той или иной ткани, органа, клетки.
7. Увеличение скорости самообновления структур, «их способности к стремительной приспособительной перестройке» как дополнительное условие лабильности компенсаторно-приспособительных реакций двух типов (свойство дуализма).
Клинический аспект современных представлений о структурных основах адаптации и компенсации нарушенных функций:
1. Всегда есть не непосредственное произведение от морфологических изменений органов, а равнодействующая от диалектического взаимодействия двух противоположно направленных биологических процессов – разрушения структур под влиянием патогенного фактора и усилий организма, направленных на ликвидацию последствий этого разрушения и восстановление гомеостаза.
2. Морфологически измененная клетка может нередко сохранять достаточно высокий уровень функциональной активности».
3. Нормализация нарушенных функций очень часто обеспечивается на основе интенсификации регенераторно гиперпластических процессов не в самом месте повреждения, а рядом или даже на отдалении от него, как возможное следствие несбалансированного перераспределения резервных возможностей, не допустимых с точки зрения грамотной подготовки и исполнения лечения практическим врачом.
4. Структурные изменения могут возникать раньше или одновременно с функциональными, но никогда не развиваются позже них (?)[19]
5. Момент клинического благополучия, т.е. вполне удовлетворительного самочувствия больного, и момент нормализации структуры пораженного органа не совпадают во времени – второй отстает от первого, причем существеннее, чем продолжительнее и тяжелее болезнь.
6. Первые клинические признаки болезни обычно не предшествуют структурным изменениям, а, наоборот, следуют за ними». Конец цитаты.
Такой доскональный и уникальный морфологический базис в структурном обеспечении адаптивно-приспособительных процессов, заложенный профессором Д.С. Сакисовым и его сотрудниками, позволяет с высочайшей степенью уверенности говорить о том, что неспецифическая компонента реакции адаптации в её долговременном исполнении (выздоровлении) имеет два типа «нормы реакции»:
1. Компенсация недостаточности функции путём гиперплазии за счёт снижения удельной нагрузки на «выздоровевшую ткань»;
2. Компенсация недостаточности функции путём увеличения скорости обменных процессов (скорости самообновления) на единицу массы (без гиперплазии).
Об этом позволяют думать и серьёзные «разночтения» в экспериментальных работах ряда исследователей, занимающихся моделированием гиперплазий и гипертрофических состояний, о которых более подробно будет представлен материал немного позже.
Поправка к теории «стресса» профессора Е.И. Чазова (1996) - «Физиология защитных реакций организма»
«Адаптационный синдром сам по себе
не является патологической реакцией;
наоборот, это необходимая физиологическая
реакция на повреждение как таковое,
имеющая защитный характер»
Ганс Селье
Не менее важными нам представляются комментарии профессора Е.И. Чазова о взаимоотношениях структуры и функции гомеостатических систем на границе нормы и патологии. Не новый, но издавна и по сей день актуальный вопрос: где грань между физиологией и патофизиологией? Это тем более необходимо для нашего изложения, ибо «Идея единого гомеостатического устройства» построена на многообразии единоподобий и всеобщности законов развития и бытия, на поиске типических неспецифических признаков и реакций, своей комбинаторикой «создающих многообразие» форм реакций (Д.С. Саркисов, см. выше).
Наверное, лучше и убедительнее сказал Клод Бернар ещё в 1866 году. Он писал: «Мы не в состоянии найти никакого коренного различия между природой явлений физиологических, патологических и терапевтических. Все эти явления происходят по законам, которые, будучи свойственны живой материи, тождественны по своей сущности и различаются только вследствие различных условий, в которых обнаруживаются явления. Физиологические законы повторяются и в патологических явлениях» (цитата из монографии Д.С. Саркисова, 1987).
Останавливаясь на этом вопросе, профессор Е.И. Чазов пишет: «Взаимоотношение структуры и функции – базис физиологии, на котором строятся все наши представления о деятельности органов и систем – представления, в конечном итоге определяющие законы существования живых организмов. Однако организм находится не в вакууме: он постоянно общается с окружающей средой, и именно взаимодействие с ней определяют законы… физиологии защитных реакций организма... Значительно хуже изучена проблематика этого раздела… Даже его название вызывает дискуссии. Почему физиологические реакции защиты, а не адаптации или компенсации? Суть в том, продолжает профессор Е.И. Чазов, что адаптация, как и компенсация, связаны с базисными физиологическими реакциями, определяющими функцию тех или иных органов либо систем. Причем адаптационные или компенсаторные возможности этих реакций поразительны…, но известен ряд специфических защитных реакций, которые предупреждают повреждение, возникновение патологического процесса (как пример: тромбообразование и синтез антикоагулянтов). Изучив эти реакции, пределы их возможностей, можно установить границы, отделяющие здоровый организм от больного». Конец цитаты.
Я бы осмелился добавить от себя: "попытаться определить разницу между понятиями «норма реакции» и «нормальная реакция»".
«С позиций практической медицины, продолжает профессор Е.И. Чазов, интересно предложение А.Н. Бруненко, рассматривающего болезнь как постоянную борьбу повреждения и физиологической защиты».
Давая определение стрессу, профессор Евгений Иванович Чазов пишет: «Стресс, - это, прежде всего, комплекс физиологических реакций на любой раздражитель из внешней среды, причем реакции связаны не только с адаптацией, но и с включением возможных (или необходимых) в данный момент защитных физиологических систем».
С этим трудно не согласиться, но чтобы в полной мере оценить возможности физиологических реакций защиты, необходимо более глубоко изучить механизмы их центральной и нейрогуморальной регуляции. Важно выяснить степень защиты и повреждения при стрессе для исследования «проблемы перехода здорового организма в больной».
Позиция очень серьезна и принципиальна: рассматривать ли адаптивно-приспособительные механизмы дифференцированно с защитными физиологическими реакциями или принять адаптивно-приспособительные реакции как средство защиты организма. Очень тяжело найти достойное возражение предложенной поправке. Она не только звучит очень убедительно, она интересна по своему существу, ибо вплотную подводит материал изложения к «Общей теории функциональных систем» профессора П.К. Анохина и принципу «Обратной связи» профессора И.И. Шмальгаузена.
Более важно иное: не искать «мнимую грань» между нормой и патологией, а попытаться установить границы резервных возможностей гомеостатических систем (клетки, организма), определяющих эту грань способностью сохранять постоянство своей внутренней среды посредством физиологических адаптивно-приспособительных (защитных) реакций.
Постулаты «Общей теории функциональных систем» профессора П.К. Анохина и профессора К.В. Судакова (1958-1996)
«Всё многообразие результатов действия функциональных систем
есть производная от функции нервной ткани, призванной историей обеспечить организм контролем и координацией эффективность гомеостатического поведения»
Автор
«Экспериментальные исследования позволили П.К. Анохину сформулировать ряд ведущих постулатов общей теории функциональных систем», - пишет его ученик профессор К.В. Судаков (1996), суммируя основные идеи построения «функциональных систем П.К. Анохина» (рисунок 8):
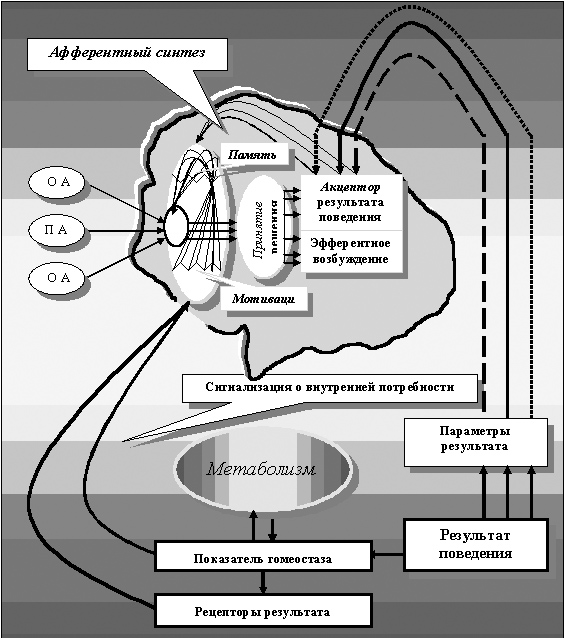
Рисунок 8. Принципиальная схема функциональной системы с её центральной архитектоникой по П.К. Анохину и К.В. Судакову
1. «Ведущим систематизирующим фактором функциональной системы любого уровня организации является полезный для жизнедеятельности организма, приспособительный результат.
2. Любая функциональная система организма строится на основе принципа саморегуляции: отклонение результата от уровня, обеспечивающего нормальную жизнедеятельность, посредством деятельности соответствующей функциональной системы само является причиной восстановления оптимального уровня этого результата.
3. Функциональные системы являются центрально периферическими образованиями, избирательно объединяющими различные органы и ткани для достижения полезных для организма приспособительных результатов.
4. Функциональные системы различного уровня характеризуются изоморфной организацией: они имеют однотипную архитектонику.
5. Отдельные элементы в функциональных системах взаимосодействуют достижению их полезных для организма результатов».
Говоря о «результатах», в постулате первом теории П.К. Анохина и К.В. Судакова, авторы выделяют 5 их типов:
1. Метаболические – «это результаты деятельности многочисленных метаболических молекулярных процессов в организме, вследствие которых образуются конечные продукты, необходимые для процессов жизнедеятельности тканей, отдельных органов и их частей или же, наоборот, - продукты нарушающие нормальную жизнедеятельность», если их наличествует избыточное или недостаточное количество «и подлежащие удалению из организма» или их накоплению, что, по существу, и составляет «норму реакции» любой гомеостатической системы.
2. Гомеостатические – «ведущие показатели крови и других жидких сред организма – лимфы, межтканевой жидкости. Гомеостатические результаты представлены в крови уровнем питательных веществ, газов, осмотического давления, рН, кровяного давления, температуры, содержания физиологически активных веществ: гормонов, олигопептидов, нейромедиаторов и т.д. Все эти показатели в их совокупности обеспечивают разные стороны нормального метаболизма тканей и составляют динамическую внутреннюю среду организма», т.е. суть метаболической «нормы реакции» в системе гомеостатической регуляции на уровне клетка – организм.
3. Результаты поведенческой деятельности животных и человека, «удовлетворяющие их ведущие метаболические, биологические потребности: специальные пищевые, половые, защитные, выделительные и другие биологические потребности, воздействующие на окружающую среду.
4. Результаты «стадной» (зоосоциальной) деятельности животных – «иерархические соотношения в популяции, общие убежища, обобщённым удовлетворением пищевых, половых и других потребностей. Деятельность животных, направленная на достижение эоосоциальных результатов нередко приобретает альтруистический характер в интересах сообщества и, прежде всего, его выживания» как свойство гомеостатически организованных участников в отношениях организм – популяция -биосфера.
5. Результаты социальной деятельности человека – «развитие учебной, производственной деятельности, бытовой активности, мероприятиям по защите общества, общением с предметами культуры, искусства и т.д. Строится специальная функциональная система, определяющая его (человека) психическую, мыслительную деятельность»[20];
6. Функциональные системы и их отдельные части избирательно созревают в процессе онтогенеза, отражая тем самым общие закономерности системогенеза.
«Центральная операционная архитектоника» функциональных систем, описываемых «Теорией функциональных систем» получила серьезное развитие учеником и последователем профессора П.К. Анохина, профессором К.В. Судаковым, подразумевая, бесспорно, функциональные системы организма человека и высших животных. Она включает в себя следующие механизмы:
1. Афферентный синтез;
2. Принятие решения;
3. Аппарат предвидения потребного результата – акцептор результата действия;
4. Формирование программы действия – эфферентный синтез, обеспечивающий реализацию принятого решения в действие, т.е. способ достижения адаптивного результата;
5. Многокомпонентное действие;
6. Сличение при помощи обратной афферентации параметров достигнутого результата в акцепторе результата действия.
По своей сути, выводы конструктивной теории профессора П.К. Анохина построены на информационном единстве механизмов управления целостной системой организма (афферентных и эфферентных процессах), как независимой гомеостатической системы, рассматриваемых с позиций классической физиологии, обобщающей и развивающей опыт своих выдающихся предшественников: И.П. Павлова, И.М. Сеченова, В.И. Вернадского.
Тем более важно, что принципы управления и самоуправления, рассматриваемые «Теорией функциональных систем» весьма применимы и на более высоком уровне организации живой материи. В нашем толковании – это гомеостатическая система планетарного уровня: биосфера Земли. Это отчетливо прослеживается в работах профессоров А.Н. Северцова, К.А. Тимирязева, Н.И. Вавилова и др. Главным объединяющим звеном такого толкования стали исследования профессора И.И. Шмальгаузена, выполненные им при работе над теорией информационного единства «всего живого».
«Теория функциональных систем» в современном её толковании также предполагает наличие общих свойств функциональных систем и более высокого уровня организации (В.К. Судаков, 1996):
1. «Устойчивость результата деятельности системы, которая достигается соответствующими механизмами саморегуляции;
2. Постоянная оценка достигнутого результата;
3. Наличие множественных исполнительных механизмов активного воздействия на результат;
4. Взаимодействие отдельных элементов системы достижения полезного для системы результата;
5. Общая функциональная архитектоника».
По мнению автора именно наличие этих свойств и этих условий определяют принцип изоморфизма любой функциональной системы[21].Так, профессор В.К. Судаков пишет:
1. «Функциональные системы гомеостатического уровня[22] могут осуществляться целиком на основе врожденных вегетативных механизмов саморегуляции;
2. Функциональные системы популяционного уровня имеют особую организацию. В них отдельные особи со своим набором функциональных систем играют роль составляющих компонентов (лидеры, исполнители, сторожи и т.д.);
3. Функциональные системы социального уровня[23], особенно производственной деятельности человека, включают взаимодействие людей не только с себе подобными, но и с определёнными техническими устройствами. В функциональной системе социального уровня для достижения значимых результатов объединяются отдельные индивиды и их функциональные системы гомеостатического уровня[24];
4. Функциональные системы психического уровня, определяющие мыслительную деятельность человека, имеют эндогенное звено саморегуляции внутренней речи, которое постоянно взаимодействует с внешним поведенческим звеном, определяющим воспроизведение устной и письменной речи и формирование внешнего движения... (В.К. Судаков, 1992, 1993);
5. Взаимодействие систем в целом организме..., имеющих следующие общие принципы:
· Принцип иерархии, развивающий принцип «доминанты» А.А. Ухтомского;
· Принцип мультипараметрического и последовательного взаимодействия..., определяющий их обобщенную деятельность;
· Доминирование функциональных систем в целом организме, определяющееся их биологической, а для человека – в первую очередь, социальной значимостью;
· Принцип системогенеза как избирательном созревании функциональных систем и их отдельных частей в процессе онтогенеза... как «квантов жизнедеятельности» или «квантов поведения» (К.В.Судаков, 1993)..., выделяющий возрастной (от рождения до старости), популяционный (формирующий отношения в стаде) системогенез, как врожденные и приобретенные «кванты» жизнедеятельности – «кванты» гомеостаза[25] (врожденные и приобретенные)». Конец цитаты.
Таким образом, «Общая теория функциональных систем» не только «заполнила» тот «пробел» в теории стресса профессора Ганса Селье, отмеченного профессором М.Г. Дурмишьяном как дефицит «роли нервных механизмов», но и существенно дополнила строительство фундамента для «Идеи единого гомеостатического устройства», системно подтвердив их роль, значение и иерархическую подчинённость, не мыслимую без существования высокоточного и оперативного функционального управления и контроля не только на уровне клетка-организм, но и на уровне организм-популяция.
Не следует только забывать о том, что всё бесчисленное многообразие результатов действия саморегулируемых функциональных систем есть суть ни что иное, как результат эволюционного усложнения живой материи, производная от функции нервной ткани, рождённой филогенезом при разделении обязанностей клеток в многоклеточной кооперации, воспроизводимой в онтогенезе из эктодермального листка и призванной самой историей обеспечить вторичный гомеостаз организма функцией совершенного контроля и координации эффективности метаболических процессов и гомеостатического поведения, т.е. управления.
Принцип «обратной связи» в едином информационном устройстве органического мира по профессору И.И. Шмальгаузену (1968)
«Человек есть, конечно, система, как и всякая другая,
подчиняющаяся неизбежным и единым для всей природы законам,
но система, в горизонте нашего современного научного видения,
единственная по Высочайшему саморегулированию...»
И.П. Павлов[26]
Имя профессора зоологии Ивана Ивановича Шмальгаузена (1884-1963) широко известно учёным не только в нашей стране, но и за рубежом. Его работы и лекции, посвящённые общим вопросам биологии развития, физиологии биологических сообществ, популяций, образования и поддержания видового постоянства, теории саморегуляции и стабилизирующего отбора в эволюции живой материи стали хрестоматийными для студентов и научных работников многих поколений, найдя своё блестящее продолжение в трудах его учеников и последователей, продолжающих систематизировать, развивать и публиковать его мысли и идеи.[27]
Достаточно вспомнить теорию «всеобщности явления регуляции в органическом мире», построенной на описании механизмов самоорганизации, заставившую по-новому взглянуть на эволюционное учение Чарльза Дарвина.
Говоря об уровнях интеграции «живого», профессор пишет: «Живые существа всегда самоорганизуются, используя внешнюю среду для восстановления и усложнения собственной организации... В основе этой саморегуляции лежат биохимические процессы. Организм строится по шаблонам, которые наследуются и размножаются». Конец цитаты.
Если эволюционное учение Ч. Дарвина открыло путь к пониманию факта возникновения, развития и многообразия видов «живого» на Земле, биогенетический закон Э. Геккеля и Ф. Мюллера позволил определить естественную модель эволюции в онтогенезе, Т.Г. Хаксли и Дж. Симпсон утвердили за организмами способность к самоорганизации, как самовоспроизводящихся систем, А.Н. Северцов, К.А. Тимирязев, В.И. Вернадский и Н.И. Вавилов сформулировали фундаментальные принципы изменчивости, наследственности и естественного отбора, то профессор И.И. Шмальгаузен своей теорией «всеобщности явления регуляции в органическом мире» объединил их представления единым механизмом управления.
Он пишет: «Самовозникновение жизни связано с интеграцией на молекулярном уровне организации... объединением аминокислот в сложной молекуле белкового вещества и синтезом нуклеидов в молекуле нуклеиновой кислоты... Однако, объединение линейной структуры нуклеиновых кислот с белковыми телами и поддержание «подвижного равновесия» в сравнительно устойчивой системе означало возникновение нового качества - возможности самовоспроизведения как основы жизненных процессов. Только путем воспроизведения создавалось всегда множество сходных индивидуальностей». Конец цитаты.
Развивая идейную основу единой системы управления на уровне организм – клетка, профессор И.И. Шмальгаузен пишет: «Интеграция и стабилизация органических систем и, прежде всего, наиболее простой системы – клетки, сопровождается в эволюции развитием компенсаторных (буферных) механизмов, которые защищают организм от вредных внешних влияний, вызывающих опасное нарушение устоявшегося равновесия». Конец цитаты.
Во истину, остается только поражаться научной смелости автора этих слов. Ведь этой фразой И.И. Шмальгаузен явно указывает на адаптивные «буферные» процессы в системе гомеостатической регуляции клетки, фактически предсказывая природу их термодинамического неравновесия, прямо называя его «подвижным равновесием».
Более того, равновесием генетически детерминированным посредством механизмов, «морфофизиологически» закрепляющих наследуемую информацию: «они (механизмы), пишет И.И. Шмальгаузен, - строятся... на регуляциях клеточного метаболизма под контролем унаследованных структур... по типу замкнутого цикла с обратной связью... как в физиологических реакциях, (позже – позиция, не только полностью подтверждённая, но и существенно развитая профессором Ф.З. Меерсоном) так и в морфогенетических процессах», изучению и блестящей систематизации которых посвятили свои научные работы профессор Д.С. Саркисов, профессор В.А. Фролов, профессор В.С. Пауков и многие другие.
Далее, развивая идею доминирующей роли «унаследованной» информации в регулировании эволюции, он продолжает: «Дифференциация, обособление частей сомы с образованием клеточных комплексов (тканей) и органов, неизменно сопровождалось соподчинением частей и их функциональных интеграций в сложном (усложняющемся) многоклеточном организме...». Далее следует не менее важный и приоритетный вывод для будущих изысканий в области развития теории адаптации: «Наличие двойной дозы гена, т.е. полигенной обусловленности наследуемой информации, а также избытка индуцирующего и реагирующего материала и множественной обусловленности формообразующих реакций оказываются средствами, защищающими и совершенствующими нормальное течение онтогенеза». Конец цитаты.
Продолжая подниматься по ступеням эволюционного ряда, профессор пишет: «Хотя интеграция достигает наивысшего уровня только в многоклеточном организме, как в целом, она всегда сопровождается и прогрессивной интеграцией молекулярных наследственных структур – в хромосомах клеточного ядра», как «памяти» эволюционных адаптаций на уровне клетки, о чём много позже будет писать профессор Р.Ю. Ильюченок[28].
При этом, автор не устает подчеркивать, что «регуляторные механизмы всего организма в целом отличаются неизмеримо большей сложностью (скорее – лучшим устройством специализаций, повышающих акцепторную способность и, за счет этого, чувствительность для более качественного анализа дифференцированной информации), чем регуляция на уровне клеточной организации», характеризуя высшую форму в организации живой материи.
Не менее поражает прямое указание на механизмы, которые будут поняты только по истечении многих десятилетий: «Все регуляторные процессы осуществляются за счёт сил, действующих внутри данной системы». Остаётся только восхищаться могучим даром научного предвидения человеком, первым среди биологов шагнувшим за грань «термодинамического неравновесия» при «избыточном производстве энтропии» в объяснениях организации живой материи на уровнях от клетки до биогеоценозов. Именно это позволило автору сделать весьма важные выводы:
1. «Биологическая регуляция – есть всегда саморегуляция;
2. Закономерное изменение структуры популяции, соответственно историческим изменениям соотношений с внешней средой, и называется эволюцией;
3. Эволюция всего вида в целом протекает в самонастраивающейся системе, состоящей из элементарных циклов регуляции каждой популяции (а также расе, подвиде) и общей регулирующей надстройки в системе вида». Конец цитаты.
Но, как ни блестящи своим содержанием работы Ивана Ивановича Шмальгаузена, при их изучении не оставляет постоянное ощущение какой-то недосказанности, внутренней неудовлетворенности автора. Его статьи, книги, очерки как бы «не завершены», не увенчаны стройной единой теорией, хотя она всегда чувствуется в общем контексте. И этот контекст как бы больше устремлён в будущее (в день сегодняшнего знания): в информатику и кибернетику.
Горько сознаваться..., но осознать всё это помогли только записи и наброски самого профессора, штрихи и комментарии так и не опубликованных его статей, некоторых глав и других материалов, ставших известными только после смерти учёного. Только ознакомившись с «Проспектом неоконченных глав» (1968), становится понятным как многому не удалось совершиться в жизни выдающегося учёного, как грандиозно много было им задумано. Так в заметках, отнесённых Иваном Ивановичем к разделу «История», он пишет: «Всегда появляются реакционные силы, не понимающие значения революций. Культ личности дал благоприятные условия. Сначала обезглавливание генетики, а затем... (события) ... 1948 года, приведшие к уничтожению подготовки молодых кадров... (эти события) отняли у нас ведущее значение в генетике, особенно популяционной, и в эволюционном учении».
Но идеализм подлинного учёного, каким был Иван Иванович Шмальгаузен, идеализм истинной науки заставляет его, с ощутимой даже читателем болью, писать: «Прогресс науки в целом остановить никому не дано. Однако все последние достижения биохимической генетики прошли мимо нас (они слишком тесно связаны с формальной генетикой – морганизмом), и теперь только у нас начинают понимать их значение». Конец цитаты.
Хочется напомнить, что это была середина XX, а не XVI столетия. Да, науку остановить нельзя, как нельзя остановить процесс познания или запретить времена года. Даже в попытках ликвидации инструментов познания и замене ценностей, нельзя отменить второй закон термодинамики и эволюцию. Жаль, что «только начав понимать», как и о чём думал великий русский учёный профессор И.И. Шмальгаузен в те 1960-е годы, «теперь у нас это» перестали делать, не умеют или/и не хотят (а ведь теперь XXI век).
В записях профессора можно найти одну удивительную по содержанию и по сути фразу, использованную им в самом широком смысле: «Научный прогресс во многом строится на аналогиях. В несколько более широком понимании, чем принято в биологии (сходство функций), они всегда использовались наукой, и это не приводит к ошибкам, если не забывать, что аналогия никогда не бывает тождеством». Конец цитаты.
Построив свою исследовательскую деятельность на доскональном анализе аналогий в устройстве «живого» на доступном тому времени уровне изучения его организации и на допустимом тем времени языке изложения, профессор И.И. Шмальгаузен уверенно соотнёс «единое информационное пространство» с «всеобщностью механизмов регуляции в органическом мире». Ему убедительно удалось показать, что явления регуляции осуществляются на всех известных уровнях организации живого: «Регуляции входят как нормальное условие функционирования всех живых систем в жизнь органоидов клетки, как целого, ткани, органа, систем органов и организма... Регуляция по принципу обратной связи присуща системам надорганизменного уровня – популяции, виду как системе и биогеоценозу...». Конец цитаты.
Даже в условиях гонений и запретов по отношению к генетике и кибернетике профессору удалось одному из первых представить эволюцию органического мира с позиций теории информации, впервые рассмотрев эволюционный процесс, как регулируемый[29].
В своей последней работе «Кибернетические вопросы биологии» (1968), вышедшей в свет уже после кончины профессора, мы находим его слова: «Управление с обратной связью включает дифференцированное участие особей в воспроизведении следующего поколения, т.е. через естественный отбор... в его преобразующей и стабилизирующей форме..., начиная с молекулярного уровня и кончая организмом как системой, до популяций и биогеоценозов». Конец цитаты.
Идеи и мысли профессора И.И. Шмальгаузена живы и успешно развиваются сегодня. Они обрели своих сторонников и последователей, сформировавших самостоятельное направление в науке, получившее название «Теории информации функциональных систем»[30]. Так в своём приветственном слове на итоговой сессии научно-исследовательского института нормальной физиологии имени П.К. Анохина, объединённого с Московской медицинской Академией имени И.М. Сеченова, принявших эстафету исследований в «Теории информации функциональных систем», профессор К.В. Судаков сказал: «Значение информационного уровня организации функциональных систем для понимания интегративных функций целого организма, роль процессов циркуляции информации состоит в обеспечении метаболического гомеостаза организма и взаимодействии субъекта с внешней средой... Представление о системе, как о любой (большой или малой) совокупности частей, деятельность которой непременно заканчивается каким-либо полезным результатом, способным воздействовать на систему и реорганизовать взаимодействие её частей не мыслимо. Без этих свойств никакое объединение компонентов не может быть названо системой».
В желании ещё более усилить бесспорное значение «информационных полей» и «информационных потоков» в составе «Теории информации функциональных систем» были предприняты две неудачные, на наш взгляд, попытки тотального обобщения введением в концепцию П.К. Анохина некоего «системогенеза», подразумевающего «избирательное созревание функциональных систем и их отдельных частей в процессе онтогенеза». Это должно было совершиться введением таких понятий и определений как:
1. «Кванты жизнедеятельности» и «кванты поведения» (В.К. Судаков, 1993);
2. Толкования информации как «всеобщего бесконечно единого законопроцесса фундаментальных отношений, связей, взаимодействий и взаимозависимостей энергии, движения, массы и антимассы микро- и макроструктур Вселенной» (И.М. Юзвишин, 1996).
По всей вероятности, неудачность попыток, в большей степени была продиктована не вполне точно избранной моделью для толкований и неполным арсеналом методологии, хотя собственно идея использования законов квантовой механики и теории информации в развитии «Теории функциональных систем» представляется более чем перспективной.
Ярким тому доказательством может служить стройная своим академизмом по сути, содержанию и форме изложения «Термодинамическая теория структуры, устойчивости и флуктуаций», развиваемая в течение многих лет профессором физики, лауреатом нобелевской премии Ильей Романовичем Пригожиным. В частности, это касается и вопросов самоорганизации «сложного»[31], упомянутых нами выше.
Основные положения теории профессора И.Р. Пригожина о термодинамической сути структуры, устойчивости, флуктуаций и самоорганизации «сложного» (1966-1994)
«Символом нашего физического мира не может быть
устойчивое и периодическое движение планет...,
этот мир неустойчивостей и флуктуаций, в конечном счете
ответственных за поразительное разнообразие и богатство форм и структур,
которое мы видим в окружающей природе»
И.Пригожин
Физик по образованию и образу мышления, лауреат Нобелевской премии профессор Илья Романович Пригожин внёс колоссальный вклад в развитие всех отраслей естествознания. Его уникальные своей универсальностью работы, закладывают надежный (трудно найти другое определение) фундамент современного знания в классические науки.
Профессору И.Р. Пригожину удалось, как нам представляется, найти подход к тому, о чём мечтали наши выдающиеся ученые: А.Н. Северцов, К.А. Тимирязев, В.И. Вернадский, Н.И. Вавилов, Н.Е. Введенский, А.А. Ухтомский, И.И. Шмальгаузен, П.К. Анохин, Г.М. Франк, Н.Н. Семенов, Л.А. Орбели, Н.М. Эмануэль и многие другие, искавшие Истину «на стыке наук».
В одной из своих работ (совместной с профессором Г. Николисом), переведенной на русский язык и опубликованной издательством «Мир» в 1990 году, - «Познание сложного», профессор И.Р. Пригожин приводит в качестве поясняющей иллюстрации тепловой самоорганизации конвекционную картину, так называемых «ячеек Бенара», возникающую на определённом этапе нагревания жидкостей (рисунок 9).
Рисунок 9. Конвекционная картина (ячейки Бенара). Размер каждой ячейки = 10-1 см;
Межмолекулярные взаимодействия = 10-8 см; одна ячейка Бенара содержит около @1021 молекул
Чтобы не нарушать логику автора и сохранить при этом избранный данной главой принцип изложения, мы также решили начать обзор основных идей профессора И.Пригожина этим примером, как это сделал сам автор. Тем более, это важно для нас, поскольку в последующих разделах этой книги мы будем часто опираться на его аксиомные мысли и обобщающие взгляды.
Возвращая изложение к ячейкам Бенара, следует особо подчеркнуть, что эта ситуация наглядно демонстрирует пример того факта, что огромное число микрочастиц, не наделенных свойством собственного гомеостаза, может демонстрировать «когерентное поведение», т.е. самоорганизовываться, не смотря на «случайное тепловое движение каждой из них», Это, по мнению автора, является «по сути одним из основных свойств, характеризующих возникновение сложного поведения».
И, как мы сможем убедиться несколько позже, профессор и в этом своём мнении не только абсолютно корректен сейчас, при описании способности микромира к самоорганизации (в конкретном примере ячеек Бенара за счет тепловой индукции), но и предельно прав «наперед». Демонстрацией «когерентного поведения» молекул жидкости сообщением тепла, можно уверенно предположить существование такого же рода организующей (управляющей) возможности для индукции другого рода: магнитной, электромагнитной, гравитационной (об этом подробнее мы будем говорить в следующих разделах).
Положив в основу своего теоретического исследования две фундаментальные теории:
1. физику неравновесных состояний «при сильных отклонениях от термодинамического равновесия» и
2. теорию динамических систем – «с превалированием неустойчивостей»,
профессор утверждает: «Существование материи связано с подавляющим доминированием частиц над античастицами, а существование жизни – с доминированием хиральных[32], асимметричных биомолекул над антиподами. Символом нашего физического мира, - продолжает свою мысль профессор, - не может быть устойчивое и периодическое движение планет, что лежит в основе классической механики. Этот мир неустойчивостей и флуктуаций, в конечном счёте ответственных за поразительное разнообразие и богатство форм и структур, которое мы видим в окружающей природе... Эта сложность присуща не только биологии. Она вторгается в физические науки и, похоже, что её корни уходят глубоко в законы природы» (конец цитаты), составляя, по существу, суть непознанного, имитирующего саму «сложность», которую подразумевает автор.
Между тем, «возможность описывать с помощью таких фундаментальных понятий как когерентность, сложность, упорядоченность, являющихся составной частью биологии, поведение как живых существ, так и обычных физических систем, является одним из основных достижений, которые, по-видимому, наука не смогла бы предсказать ещё несколько лет тому назад». Конец цитаты.
Используя ту же модель ячеек Бенара, профессор И.Р. Пригожин проводит ещё одну удивительную своим обобщающим эффектом аналогию. Зная строгую «детерминированность и повторяемость эффекта при абсолютной непредсказуемости его лево и право вращения, - профессор заключает, что – таким образом, мы приходим к удивительному сотрудничеству между случайностью и определенностью, наводящему на мысли об аналогичном дуализме, известном в биологии со времен Дарвина (мутация – естественный отбор). В области физики такой дуализм наблюдается лишь при квантово-механическом описании явлений». Конец цитаты.
Это и понятно, так как свойством дуализма наделены, со всей очевидностью, лишь «счастливые обладатели» собственного гомеостаза: органелла, клетка, организм, планетарная биосфера и, как это будет видно из последующих глав, двуединая бесконечность Вселенной и элементарные частицы.
«Таким образом, - продолжает автор, - мы видим, что вдали от равновесия, т.е. при существенном ограничении степеней свободы, система может приспосабливаться к своему окружению несколькими различными способами». Конец цитаты.
Позволим себе повторить только что высказанную мысль чуть иначе, добавив несколько уточняющих слов, не меняющих её существа: «Таким образом, мы видим, что в мнимой дали от всеобщего равновесия, т.е. при существенном детерминировании процессов областью центральной, локальной и/или частной систем управления, ограничивается их степень свободы и система может приспосабливаться к своему окружению несколькими различными способами».
В такой транскрипции, повторюсь, не меняющей сути сказанного, это определение подпадает под утверждение всеобщности механизмов адаптации, мутаций и управления. А уже как следствие этого: свойство дуализма, присущее живой и неживой материи. Кстати, о свойствах дуализма неживой материи пишет не только профессор И.Р. Пригожин, в особенности это касается элементарных её участников – элементарных частиц. Об этом более подробно мы будем говорить в разделе втором: «Участники гомеостатического устройства».
«Тот факт, - продолжает профессор, - что из многих возможных вариантов был выбран лишь один (для каждого конкретного вида многообразий, но в рамках единого устройства в управлении), придает системе историческую размерность. Своего рода «память» (о которой пишет и профессор Р.Ю. Ильюченок) о прошлом событии (закрепленном генетически в онтогенезе), произошедшем в критический момент (или момент статистической случайности) и оказавшем (оказывающем) влияние на дальнейшую эволюцию» через механизмы адаптации (срочной, долговременной, «системного структурного следа») путём перераспределения внутренних статей расходования энергии, компенсирующих локальный «дефицит энергии»[33] Конец цитаты.
«Что же происходит в том случае, когда тепловое ограничение степеней свободы превысит первый порог структурированию?» – задает вопрос автор в поисках теоретического обоснования «пороговой чувствительности» системы. И дает ответ: «В некоторой области DT ячейки Бенара будут существовать как таковые, однако некоторые их характеристики изменятся. Затем структура течений станет размытой и возникнет новый режим, характеризуемый неупорядоченной зависимостью переменных от времени... – Турбулентность – это лишь один из аспектов общей тенденции целого класса систем к переходу в хаотический режим». Конец цитаты.
Но ведь при таком толковании турбулентности, нельзя исключить, что она (турбулентность) – это своего рода «предельная», критическая фаза возбуждения, характеризуемая абсолютным преобладанием возбуждающих действий (вещества, энергии, информации) над силами структурирующими (примитивную или любую другую) систему.
При этом, «ячейки Бенара, - указывает автор, - предмет модель, в которой химическая природа вещества, образующего жидкий слой не изменяется». Конец цитаты.
Это, на наш взгляд, ещё один аргумент в пользу атрибута управления и абсолютного соответствия его условиям сохранения механизмов самоорганизации, не изменяющих качество участников в рамках сохранения их «ограниченной независимости». Т.е. демонстрация управления и независимости в борьбе за единство противоположностей в естественном исполнении и на уровне «неживого» микромира.
В случае «живой материи» описанные процессы перераспределения не могут не найти должной поддержки систем более высокого уровня организации (учитывая условия их информированности и эффективности управления), хотя эта поддержка и должна быть заключена в рамки целесообразной для них независимости (компетентности) и в полном соответствии их возможностям. Это и составляет суть классификации и суть дуализма резервов независимых гомеостатических систем:
1. через перераспределение статей расходования энергии подчинённых участников, входящих в данную систему и
2. путём привлечения гомеостатических систем эквипотенциального уровня через перераспределение их функций, управляемых вышестоящим иерархом по принципу обратной связи.
Во втором случае, ограничения резервов устанавливаются собственно обладателем гомеостаза, названного выше «иерархом более высокого уровня организации», в рамках его же внутренних возможностей и установленной им же термодинамической целесообразности.
Очень важно, что профессор И.Р. Пригожин через свойство самоорганизации находит и вычисляет механизм, посредством которого система (даже на элементарном уровне) реализует свою способность иметь не одно, а, минимум, два решения. Это – эффект «турбулентности» («флуктуаций, существующих всегда») или иначе, - «эффект возмущения», устанавливающий термодинамически выгодные условия для принятия «устойчивого» решения минимум двух типов. Это: «бифуркации при нарушении симметрии асимптотических систем» (рисунок 10). Именно таким образом объясняется механизм управляющего и самоуправляющего устойчивого действия.
Рисунок 10. Диаграмма бифуркации, показывающая влияние изменения управляющего параметра на переменную состояния X (верхняя часть рисунка);
Диаграмма последовательных бифуркаций (нижняя часть рисунка), где А и А’ – точки первичных бифуркаций из термодинамической ветви В и В’ – точки вторичных бифуркаций
![]()
Рассматривая теорию точек бифуркации (или «флуктуаций») для асимптотических (возбудимых) систем, следует, на наш взгляд, серьезно подумать об аналогии (или абсолютной идентичности) с любыми проявлениями дуализма, как доминантного фактора естественных адаптивно приспособительных регуляций гомеостатических систем.
Обсуждая далее эффект ячеек Бенара, профессор И.Р. Пригожин пишет: «конвекцию Бенара можно представлять себе следующим образом: слабые конвекционные токи, возникающие как флуктуации относительно среднего состояния, существуют всегда, но ниже некоторого критического значения градиента температуры эти флуктуации затихают и исчезают. Если же мы, - продолжает профессор, превышаем критические градиенты температуры, то некоторые флуктуации усиливаются и порождают макроскопическое течение. Возникает новый молекулярный порядок, по существу, гигантские флуктуации, стабилизируемые за счёт обмена энергией с внешним миром. Этот порядок характеризуется возникновением того, что принято называть «диссипативными структурами», приводит к весьма неожиданным явлениям, в том числе к возникновению порядка через флуктуации, взаимодействие между... тремя аспектами:
Функция ¬ ® Структура
¯ ¯
Флуктуации
Конец цитаты.
В любом контексте, применительно ли к модельной системе «надэлементарных систем» в случае ячеек Бенара, каталитическому химическому процессу, самовоспроизведению при генных репликациях или сложных многокомпонентных участников адаптивных процессов гомеостатических систем, во всех случаях действует механизм «случайной определенности», где «случайность выступает в роли некоторого инновационного элемента, нужного для прощупывания пространства состояний, а упорядоченность позволяет системе поддерживать коллективный режим (особенно это важно для управления), охватывающий макроскопические пространственные области и временные интервалы. Необходимой посылкой всех этих явлений служит нелинейная динамика, при определенных условиях приводящая к неустойчивости движения и бифуркациям». Конец цитаты.
Иными словами, во всех случаях действует «правило дуализма», которое мы позволили себе назвать правилом Пригожина: Процесс адаптации при возбуждении асимптотических систем имеет не менее двух термодинамически равноценных типов решений.
В общем плане нашего изложения, из вышеизложенного может следовать очень важное заключение: если теоретические доказательства профессора И.Р. Пригожина верны и носят общий и/или всеобщий характер, то процессу адаптации любой системы, наделенной свойством гомеостаза (частный случай асимптотического), к изменениям условий в обмене веществом, энергией и/или информацией, должно быть применимо «правило Пригожина».
Иными словами, развивая эту нить рассуждений и в полном соответствии сформулированному «правилу Пригожина»: любые гомеостатические системы должны быть привержены и/или обладать свойством дуализма. Проявлений же избранного дуализмом решения следует ожидать в период «возмущения» или возбуждения («стадия срочной адаптации») гомеостатической системы, а её морфологического эффекта - на стадии «системного структурного следа»[34].
И действительно, придя к такому заключению, становятся понятны многие необъяснимые прежде факты, полученные экспериментальным путем:
Пример 8. Так, исследуя особенности динамики ATP, АDР, АМР, неорганического фосфора и энергетического потенциала миокарда у исходно повреждённых сердец (с искусственно вызванной гипертрофией рабочего миокарда), нам долгое время не удавалось добиться стабильности цифр по результатам исследований.
Прежде всего, мы обратили внимание на то, сердца крыс, полученные после стеноза брюшной аорты и задней полой вены, либо имели существенно увеличенную массу, либо не изменяли её вовсе. Причём соотношение таких результатов было приблизительно одинаковым. Это очень затрудняло и анализ и интерпретацию.
На первом этапе, мы исследовали все сердца, не зависимо от их веса после операции. Однако, очень большая дисперсия абсолютных данных после пересчёта показателей на единицу массы при статистической обработке, не позволяла работать в рамках хотя бы условной достоверности. Но интересно, что индивидуальная динамика всегда четко повторялась от эксперимента к эксперименту.
Долгое время мы пытались найти причину в неадекватности избранной модели. Безуспешно. Затем, использовали для последующих исследований только сердца, масса которых была увеличена после операции, не обрабатывая данные «неудачных» результатов операций (сердца с неизменённой массой), создавая таким образом как бы «искусственный фильтр» для исходного материала. Но нас постоянно мучило чувство «недостаточной» корректности. Некой «подгонки» результатов.
В попытке «обойти» возникшие сложности, было принято, по тем временам, единственно возможное решение: использовать при анализе динамики относительные единицы – энергетический потенциал сердца, выраженный через массовые отношения всех участников энергетического метаболизма. Дисперсия показателей сразу стала маленькой, а результаты, поддающимися анализу.
Много позже, мы нашли прекрасное объяснение произошедшему в работах профессора Д.С. Саркисова, установившего, как мы уже отмечали, что неспецифическая компонента реакции адаптации в её долговременном исполнении (выздоровлении) имеет два типа «нормы реакции»:
1. Компенсация недостаточности функции путём гиперплазии за счёт снижения удельной нагрузки на «выздоровевшую ткань»;
2. Компенсация недостаточности функции путём увеличения скорости обменных процессов (скорости самообновления) на единицу массы (без гиперплазии).
Однако, это было «качественное» решение вопроса с позиций нормальной и патологической физиологии, не имеющего серьезного теоретического фундамента. Еще тогда мы предположили и попытались на основании анализа эмпирического материала трактовать наблюдаемый эффект регуляции адаптивных систем путем изменения скорости энергопродукции, достигаемый за счет снижения «критического» содержания АТФ (А.А. Лубяко и соавт, 1987, 1988, 1991).
Теория бифуркаций профессора И.Р. Пригожина и, как следствие, «правило дуализма» для асимптотических систем позволяют с уверенностью говорить не только о том, что наше изначальное предположение было верно, но и утверждать, что механизм регуляции адаптивных систем, обладая свойством дуализма, способен компенсировать дефицит энергетического баланса через гиперфункцию миокарда двумя средствами:
1. Компенсацией недостаточности функции путем гиперплазии и снижения удельной нагрузки на «выздоровевшую ткань» за счет уменьшения удельной энергопродукции.
2. Компенсацией недостаточности функции путем увеличения скорости обменных процессов (скорости самообновления) на единицу массы (без гиперплазии) за счет увеличения удельной энергопродукции.
Таким образом, можно было сделать и был сделан очень важный вывод: генеральным регулирующим звеном в механизме адаптации гомеостатической системы клеток сердца к условиям гиперфункционирования является удельная эффективность энергетического метаболизма.
Теперь, на базе вышеизложенного, очень важно было бы найти ответ на следующий вопрос: соответствует ли термодинамическая равноценность решений физиологической их равноценности и/или равной эффективности?
Хочется выразить предположение, основанное на выводах только что приведенного частного случая из собственного опыта: поскольку законы всеобщности (если они являются таковыми) должны быть приложимы ко всем, избранным нами в концептуальном плане носителям гомеостаза, коим является клетка, организм и т.д., то «удельная эффективность энергообеспечения» должна быть универсальным регулятором адаптивных процессов для гомеостаза любого уровня организации материи (если «Идея единого гомеостатического устройства» верна в принципе)[35].
В завершении этого, далеко не полного, но очень важного для понимания «Идеи единого гомеостатического устройства», изложения некоторых элементов теории профессора Ильи Романовича Пригожина, хочется привести его слова, сказанные во «Введении» его статьи «Единство физических законов и уровней описания». В названной работе профессор пишет: «Довольно примечательным совпадением является то, что эволюционная идея возникла в XIX веке в двух прямо противоположных формах.
· В термодинамике принцип Карно-Клаузиуса формулируется как эволюционный закон непрерывной дезорганизации, или разрушения изначально заданной структуры.
· В биологии или социологии идея эволюции, напротив, ассоциируется с усложнением организации.
Ясно, что обе концепции имеют глубокий философский смысл. Распространение термодинамической концепции эволюции на мир в целом приводит к идее о том, что «структура» возникла в каком-то весьма от нас удалённом «золотом веке» и что последующее развитие связано с непрерывным переходом к хаосу.
Биологическая концепция эволюции утверждает нечто прямо противоположное. Никто не высказал её лучше, чем французский философ Анри Бергсон, взгляды которого отражают состояние биологии того времени: «Чем глубже мы проникаем в природу времени, тем больше понимаем, что длительность есть изобретение, создание форм, непрерывное производство чего-то абсолютно нового. Идеи Спенсера в некотором смысле аналогичны. Спенсер полагает, что основным принципом эволюции в природе служит принцип «неустойчивости однородного». Конец цитаты.
«Как можно примирить эти два диаметрально противоположных аспекта эволюции?» - задает вопрос профессор И.Р. Пригожин.
Нам представляется, что «примирение» не только возможно, но и неизбежно, если путь к нему проложен такими учёными как профессор И.Р. Пригожин, профессора E. Pfluger, L. Fredericq, Н.Е. Введенский, C.Bernar, А.А. Ухтомский, И.П. Павлов, W.Cannon, А.Сперанский, А.Н. Северцов, К.А. Тимирязев, В.И. Вернадский, Н.И. Вавилов, H. Dotterweich, Г.М. Франк, Л.А. Орбели, Н.М. Эмануэль, К.М. Быков, П.К. Анохин, H.Selye, В.В. Парин, Ф.З. Меерсон, И.И. Шмальгаузен, В.А. Фролов, Р.Ю. Ильюченок, Н.А. Онищенко, Д.С. Саркисов, К.В. Судаков.
И эта неизбежность тем более «примирительна», что путь к ней лежит через всё и всех объединяющую «Теорию информации», рождённую XX веком и всё более завоёвывающую мировое «информационное пространство» на пороге нового тысячелетия.
Определения и суть определений «Теории информации и информационных пространств»
«Чем больше круг наших знаний, тем большая поверхность его
соприкосается с незнанием»
Платон (428/427-348/347 г до н.э.)
По существу, суть теории информации может быть сведена к положениям, корректно отвечающим на следующие основные вопросы: Что есть информация или что может быть названо информацией как таковой? Каковы способы ее приема? Как эта информация может быть передана? Где и каким способом эта информация может быть сохранена? Что такое память?
Вот, собственно, весь круг вопросов, ответом на которые смог бы удовлетвориться пытливый ум исследователя, не пытающийся усложнить свою и без того нелёгкую свою творческую жизнь. Ибо найдя ответы на эти вопросы, можно было бы установить и способы измерения информации, и принципы контроля, и методы накопления, оценить качество.
В принципе, в том виде, в котором существует теория информации сегодня, это ветвь теории вероятностей и математической статистики. Поэтому абстрактные формулировки могли бы стать применимы к любой вероятностной или статистической системе наблюдений. И действительно, теорию информации можно использовать практически во всех областях знания, так же как теорию вероятностей и статистику.
Что же касается духа и идей теории информации, то они «вырастают из понятия беспорядка или энтропии в термодинамике и статистической механике» (Фишер 1935, Шеннон, Уивер 1949, С. Кульбак 1967). Главными операторами этой теории служат такие понятия как информация, количество информации, качество информации, память.
С точки зрения философии, информация[36] – это разъяснение, изложение или некоторые сведения, совокупность каких-либо данных, знаний, являющихся одновременно одним из основных понятий кибернетики (А.В. Адо и соавт. 1986).
Само рождение «Теории информации» сыграло в науке, в особенности в прикладных её дисциплинах, роль, аналогичную понятию энергии в физике, также давшему возможность с обобщающих позиций подойти к описанию самых различных физических процессов, пытаясь проникнуть глубже в их суть.
Особенно ключевую роль она играет в теории связи, рассматривающей собственно систему связи как стохастический, или случайный, процесс. Так Таллер (1950) отмечает, что статистическую теорию связи часто называют теорией информации. Ротштайн (1951) даёт определение теории информации как «чистой механики, оперирующей измеримыми множествами, с выбором из альтернатив неопределённого характера». Пирс (1956) считает, что теория связи и теория информации – одно и то же. Гильберт же (1958) утверждает: «Информация – это такая мера времени или стоимости, которая, в частности, нужна инженеру при планировании эксперимента». Шеннон (1949, 1956), Фишер (1956) и Виннер (1956) рассматривают информацию как «сведения, выраженные условными знаками об объекте или его стороне». По Харкевичу «информация - это степень изменения вероятности достижения цели». Школой информатики Технического Университета им. Н.Э.Баумана (Москва) и Московского инженерно физического института информацию принято квалифицировать «как количество простых фактов, называемых SEM, где каждый простой факт содержится в минимальном неделимом предложении» (В.В. Воробьев, Китов, 1989).
По существу же, математический и статистический характер теории информации всегда оставался неоспоримым.
По этому поводу «существует обширная литература», пишет в своей книге «Теория информации и статистика» профессор математики Соломон Кульбак (1968), посвящая её «изучению соотношений между понятиями и математической формой энтропии и информации» со ссылкой на. стр. 8-11 библиографической ссылки профессора Стюмпреса (1953) и приводя свой, не менее обширный перечень литературных источников, начиная с 1928 года, с работ профессоров Хартли, Черри (1957) и Фрезера (1957) и заканчивая трудами наших современников (стр. 11-13).
Не вступая в противоречие мнению большинства учёных, в попытке не утяжелять и без того сложный предмет изложения, мы можем позволить себе квалифицировать теорию информации как «стилизованную символами логики (квантерами) математическую статистику».
Исходя же из того, что собственно математика «есть суть инструмент» любой науки, можно позволить себе рассмотрение «Теории информации и информационных пространств», ограничившись анализом основных её определений, положений и тезисов, составляющих базу обширнейшего материала.
В этой части описания мы, конечно, больше внимания уделим теории информации, исходя из тех задач и тех позиций, которые продиктованы существом «Идеи единого гомеостатического устройства», где одним из основополагающих принципов является поиск способов объективной оценки функционального состояния гомеостатических систем путём качественного и количественного измерения процессов обмена веществом, энергией и информацией на всех уровнях организации живой материи между её участниками.
Первым из естествоиспытателей биологов, кто рассмотрел теорию эволюции органического мира с позиций информатики, был профессор И.И. Шмальгаузен. Он впервые применил математический инструмент как некую меру при описании многообразия форм, оценив её количеством информации, измеряемой суммой отрицательных логарифмов вероятностей всех вариантов X форм, составляющих анализируемое многообразие[37], где:
X = - å log pi, а величина «средней» информации (I) равна: (1)
I = -p1 log p1 – p2 log p2 – p3 log p3 = - å pi log pi (2)
Это "скромное" математическое выражение (Фишер 1935, Шенон 1949, 1956, С. Кульбак 1967 и др.) стало классическим для собственно «Теории информации», применимой, в частности, к биологическим системам (И.И. Шмальгаузен 1968, И.А. Шилов 1985, В.К. Судаков 1996, И.И. Юзвишин 1996, Л.П. Агулова с соавт. 1998 и другие).
Это тем более важно для нашего предмета изложения, поскольку хотелось бы особо акцентировать внимание на том факте, что при количественной оценке, информация рассматривается всеми авторами как величина с противоположным знаком по отношению к среднему вероятностному событию.
При этом, представление об информации отвлекается от содержательной стороны сообщений и рассматривает только её количественный аспект. Так и появилось понятие «количество информации» I(x) - величина, обратная вероятности того события, о котором начинает идти речь при анализе какой-либо системы (А.В. Адо и соавт, 1986).
Иными словами, если X – событие, а его распределение нормально и имеет параметры a и s согласно закону нормального распределения Гауса N(x; a, s), то:
j(x) = e-x/2 (1/Ö2p) (a=0;s=1), (3)
информация же, представляя собой по определению величину, обратную вероятности события, правомерно записать как:
_____
I(x) = 1/j(x) = e-x/2 Ö2p (4)
Из этого следует что, чем более вероятно событие X, представленное функцией j(x), тем меньше информации I(x) несёт анализируемое сообщение о его наступлении, и наоборот[38]. Это заключение полностью корректно философскому толкованию информации, представленному выше (А.В.Адо и соавт, 1986).
Поскольку известно, что в понятии информация следует различать два аспекта:
1. Информация I(x) представляет собой меру организации системы.
В этом случае, её математическое выражение тождественно выражению для энтальпии, но, если энтропия системы (S):
S = (U + pV)/ T , то энтальпия системы (H) (5)
H = U + pV , т.е. количество информации (I) (6)
I = U + pV (7)
где: U – энергия, H – энтальпия, S – энтропия, p - давление, V- объем, T – температура.
Из приведённых выражений видно, что как энтропия системы выражает степень её неупорядоченности, так и информация указывает меру её организации.
2. Понятая (принятая и/или акцептованная) информация составляет внутреннее достояние системы и/или процесса и должна быть названа (становится) структурной информацией, т.е. входит элементом в структуру «памяти»
Теория информации, опираясь на аппарат математической статистики и второе начало термодинамики, позволяет исчислять размерность количества информации, выраженной в безразмерных единицах – битах, в физических единицах энтропии (Дж/К) (см. Главу 4).
На основании изложенного, и информацию и энтальпию, в полном соответствии выше сформулированному определению, можно анализировать как противомножество от события, произошедшего и/или происходящего в этой системе[39].
По сути же, следующей из вышеприведённых определений авторов, представляющих большинство отраслей современного знания, информацией можно назвать всё то, что может быть акцептовано какой-либо системой.
В связи с тем, что по определению информация есть суть величина термодинамического свойства, то её акцепт должен сопровождаться изменением установившегося баланса в обмене веществом и энергией.
В свою очередь, изменение в обмене веществом и энергией может быть квалифицировано «теорией возмущений» (М. Дьюар, Р. Догерти, 1977), как реакция возмущения, что полностью соответствует «принципу обратной связи» И.И. Шмальгаузена.
Согласно же квалификациям «Теории функциональных систем» П.К. Анохина и К.В. Судакова такое свойство является характеристикой организма, т.е. организованной в высшей степени материи, - живой материи.
Таким образом, с точки зрения «Идеи гомеостатического устройства» информацией можно назвать всё то, что связано со способностью «участников» обмена веществом и энергией, принять (акцептовать) информацию и модулировать её до уровня дееспособной, т.е. способной вызвать специфический возвратный физиологический ответ.
Если же подходить к вопросу по-философски шире, то наверное, на основании всего изложенного выше, и помня о необходимости ответить на вопрос этой главы: «что же такое информационное пространство», можно сказать так: информационное пространство – это театр любого действия в мироздании, где единой мерой и универсальным инструментом его регулируемого качества является энергия.
Комментарии к основополагающим учениям «Идеи единого гомеостатического устройства» (вместо заключения по материалам главы)
В плане общего построения нашего изложения и собственно предпосылок к формулированию основных положений «Идеи единого гомеостатического устройства», представляется чрезвычайно важным безотносительный аналитический подход выше приведенных академических теорий, эмпирических предположений, классических учений и постулатов.
Поставив своей целью выделить на базе существующих знаний о «частном» общие и всеобщие черты, принципы, закономерности и законы, мы умышленно не акцентировали внимание на более мелких частностях отдельных биохимических и других реакций и процессов. О них мы будем говорить ниже, в последующих главах этой книги. Сейчас же, мы пытались найти, говоря языком физиологов «неспецифические», «типические» компоненты общего построения, общего обустройства, общих принципов управления и реакций в обобщённом понимании структуры, функции, механизмов действий и взаимодействия.
В заключение этой главы, хочется ещё раз процитировать очень важные для нашей идеи, опережающие современное знание слова одного из выдающихся учёных, блистательного врача-эмпирика нашего времени профессора Ганса Селье: «…принципы защиты на уровне клетки в основном приложимы и к человеку и даже к целым сообществам людей» (1979).
От себя готов добавить: "эти принципы приложимы на всех уровнях организации «живой» материи, на всех уровнях её единого гомеостатического устройства".
Глава 4
Всеобщие, общие и частные законы,
«Идеи единого гомеостатического устройства»
[1] «Интервью в парижском такси», 1933 год
[2] В качестве ремарки: само по себе весьма примечательно, что «эволюция» - это «слово-мутант», изменившее во времени свое исходное назначение от «военного, тактического и стратегического движения армии и флота» (В. Даль, 1882) до «процесса постепенного непрерывного количественного изменения, подготавливающего качественные изменения; или вообще - развитие во 2 значении» (С.И. Ожогов, Н.Ю. Шведова, 1999), трактуемое современной философией как «изменение бытия и сознания» (И.Т. Фролов, 1986).
[3] от пара...- приставка, обозначающая: прилежащий или расположенный вблизи чего-либо или напоминающий что-либо, или аномальный и ...биоз – приставка, указывающая на связь с жизнью (W.Alexander et al. 1999)
[4] Автор пишет о своём учителе профессоре Н.Е. Введенском
[5] «нормальных органных выделений» по J. Dausset (1959)
[6] Среди незаслуженно забытых имён, прямо указывавших на этот сложный в композиционном и крайне важный в физиологическом смысле агент, следует назвать профессора Черткова и доктора А. Фриденштейна (из выступления профессора В.И. Смирнова на XIII (76) сессии общего собрания Российской Академии медицинских наук 17-20 февраля 2004 г).
[7] Ремарка: профессор А.А.Ухтомский (опубл. в 1950 году), который всегда был и останется высочайшим примером и образцом научной корректности для всех нас и наших коллег, писал о своем учителе: «Если бы он удовлетворился тем, что им сделано до 1886 года, и стал лишь логически обрабатывать, подчищать и популяризировать свои тогдашние взгляды, его имя было бы чрезвычайно популярно в современной нам науке, которая именно в последние годы утвердилась на его тогдашней точке зрения».
· Будучи известным всему миру учёным, удостоенным бесчисленных званий и наград, профессор Алексей Алексеевич Ухтомский не переставал говорить о том, что «… школа, к которой я принадлежу, школа профессора Введенского…, а «доминанта» есть естественное и неотъемлемое детище школы Введенского». (В собр. соч. 1950 г., с.232-292).
· Великий ученый, академик И.П. Павлов сказал однажды по этому поводу: «При изучении этих отклонений (имея ввиду ЦНС) в сторону преобладания торможения, ослабления раздражительного процесса, нам пришлось убедиться, что одно из открытий нашего выдающегося покойного физиолога Н.Е.Введенского глубоко справедливо. Введенский сделал очень много в нервной физиологии, ему посчастливилось найти крупные факты... он устанавливает изменения нервного волокна под влиянием сильных раздражителей и различает при этом несколько фаз. И вот оказывается, что эти своеобразные фазы целиком воспроизводятся и на нервных клетках, когда вы сильно напрягаете борьбу между раздражительным и тормозным процессами» (И.П.Павлов, Полн. Собр. Соч. т.3, кн.2, стр.28)
· Если присмотреться внимательней, то в этом высказывании всемирно авторитетного учёного можно найти обозначение почти всех направлений, которыми сегодня занимаются наши современники, пытаясь подойти к их разрешению, которые не только не потеряли своей актуальности, но, напротив, всё более её приобретают.
· Не даром же профессор А.А.Ухтомский пишет горькую, но такую традиционную для нашего отечества правду: «Покойного ценили при жизни очень мало, в особенности в России. Сам же он не делал никаких шагов к тому, чтобы обратить на себя внимание или популяризовать свои взгляды».
[8] официальная резолюция цензора
[9] М.Е. Райскина, Н.А. Онищенко, Б.М. Шаргородский, К.М. Халимова, Б.Н. Фельд, Б.П. Расторгуев. Методы прижизненного исследования сердца, М, Медицина, 1970, 264 с.
[10] в то время ещё кандидата наук, старшего научного сотрудника
[11] Шумаков В.И. Механизмы адаптации трансплантированных органов. В журн: Вестник РАН, 1996, том 66, N 12, с.1072-1078
[12] Лубяко А.А. Механизмы формирования и принципы диагностики противоишемической резистентности миокарда. Дис. ... Докт. биол. наук, М.: 1994, 487 с.
[13] Лубяко А.А., Онищенко Н.А., Курапеев И.С., Новиков В.К., Ефременков Е.А. Применение кардиоплегических растворов в кардиохирургии. Методические рекомендации. Л, 1988, 19 с.
[14] Новиков В.К., Курапеев И.С., Лубяко А.А., Николаева И.П., Лобков А.В., Ефременков Е.А., Поликарпов И.С., Гайдова О.С. Противоишемическая подготовка кардиохирургических больных витамином Е при операциях на "открытом сердце"./Методические рекомендации. Л.: 1990, 15 с.; Новиков В.К., Ефременков Е.А., Гороховатский Ю.И. и др./Доклад на заседании Ленинградского научного кардиологического общества им. Г.Ф.Ланга 27 февраля 1984 г., Л, 1984; Новиков В.К., Головин В.Н., Котин А.Н., Шнейдер Ю.А., Курапеев И.С., Лубяко А.А., Ефременков Е.А., Поликарпов И.С., Фёдотов А.Г., Лобков А.В. Успешное хирургическое лечение травматической недостаточности митрального клапана (падение с высоты 10 метров)/Доклад на заседании Ленинградского научного общества имени Г.Ф.Ланга 12 декабря 1988 г с демонстрацией. Л, 1988; Новиков В.К., Лубяко А.А., Александров К.Ю. Развитие клинической физиологии в кардиохирургии./ В журн.: Грудная хирургия. 1989, 6, с.13-16; Новиков В.К., Курапеев И.С., Ефременков Е.А., Лубяко А.А. и др. Разработать и передать в практику способ противоишемической защиты миокарда у больных с ишемической болезнью сердца и при кардиохирургических вмешательствах с использованием диагностических приборов, средств и методов фармакологического воздействия./Отчет о НИР Инв.N 02910026329, Л:1990, 62 с.
[15] Более того, опытный перфузиолог, как показали исследования, интуитивно нарушает канонические режимы перфузии, интуитивно ощущая реальную потребность ответные противодействия организма, невольно «попадая» в зону, единственно оптимальную для данного пациента. Однако, это ни в коей мере не гарантирует его от ошибок практически в каждой проанализированной операции.
[16] Лубяко А.А., Курапеев И.С. Энергобаланс сердечно-сосудистой системы во время искусственного кровообращения./В журн: Грудная и сердечно-сосудистая хирургия, 1994, No5, с.34-36
[17] Разница между суммарным (∑) количеством электронов (N), способных разместиться на электронных оболочках (n) атома при полном их заполнении (∑Nn при Nn = 2n2, n=1,2,3,4,5,6,7) и реальным количеством электронов атома данного вещества.
[18] Ремарка профессора В.А.Фролова: «При K=1 в сердце имеет место соответствие выработки энергии потребности в ней миокарда. При K<1 возникает энергетический дефицит, если K стремится к 0, речь идёт о прогрессирующем истощении сердечной мышцы.
[19] Это довольно спорное заключение, поскольку факт изначальных структурных изменений всегда возможен в этиологии целого ряда заболеваний, но его решающая в обобщении этиологическая роль не может являться, на наш взгляд, абсолютной.
[20] по Ф.Энгельсу: «Бытие определяет сознание»
[21] С этим положением сложно согласиться, прибывая в рамках установления не только общностей, но и всеобщности. Именно поэтому нижеприведенные аргументы, на наш взгляд, могут быть рассмотрены как проявления общего на уровне частного устройства. Тем не менее, они имеют важное значение и в плане нашего изложения.
[22] Исторически подразумевается уровень организма
[23] Весьма спорное определение в классификации биологических систем. Скорее оно носит характер классификации психологического, даже, социологического свойства.
[24] Это положение, а равно следующее чрезвычайно важно, ибо позволяют впервые создать условия наибольшего благоприятствования для экстраполяции знаний академических и естественных дисциплин в область психологии, живущей, в определенном смысле, на значительной дистанции от физиологии субъекта их исследований. И наоборот, по «принципу обратной связи», физиологам приобрести опыт, накопленный психологами.
[25] Мы не будем останавливаться на комментариях к этим «не понятым сегодня» принципам, но считали корректным их упомянуть в «авторском исполнении».
[26] Павлов И.П. Ответ физиолога психологам \Полное собрание сочинений\\2-е издание. М.1951, т.3, ч.2, с.153-188
[27]Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М.1939; Шмальгаузен Кибернетические вопросы биологии. Новорсибирск.1968; Шмальгаузен И.И. Факторы эволюции. Теория стабилизирующего отбора. М.1968; Шмальгаузен И.И. Проблема дарвинизма. Л.1969, 2-ое издание; Шмальгаузен И.И. Организм как целое в индивидуальном и историческом развитии. М.1982; Шмальгаузен И.И. Пути и закономерности эволюционного процесса. Избранные труды. М.1983
[28] Ильюченок Р.Ю. Память и адаптация. Новосибирск, Наука, Сиб. Отделение, 1979, 192 с.; Ильюченок Р.Ю., Дубровина Н.И., Подгорная Е.К. Механизмы эмоциогенной регуляции памяти \Вестник РАМН, 1998, 9, с.24-29
[29] Чем это не пример резервных возможностей живой развивающейся материи.
[30] Им «не научившимся и/или не умеющим понимать» так и не удалось даже «отстать» нас от наших же западных коллег.
[31] Пригожин И., Дефей Р. Химическая термодинамика. Новосибирск, Наука, 1966, 509 с.; Николис Г., Пригожин И. Познание сложного. М.Мир, 1990, 342 с.; Гленсдорф П., Пригожин И. Термодинамическая теория структуры, устойчивости и флуктуаций. М.Мир, 1973, 280 с.; Пригожин И. Порядок из хаоса: новый диалог человека с природой. М.Прогресс, 1986; Пригожин И. От существующего к возникающему. Время и сложность в физических науках.. М.Мир, 1985, 327 с.; Пригожин И. Время, хаос, квант. М.Прогресс, 1994
[32] от греческого – cheir – рука, т.е. принадлежность объекта к одной из зеркально равных – «правой» или «левой» модификации
[33] Мы позволяем себе комментарии «в тексте» авторского высказывания, в виде фраз, заключённых в скобки, авторский же текст везде выполнен нормальным шрифтом и сохранен дословно.
[34] В соответствии с классификаций Ф.З. Меерсона
[35] К этому вопросу мы будем обращаться во всех последующих главах, оперируя такими понятиями как «удельная эффективность функционирования», «удельная эффективность энергообеспечения», «удельная скорость энергообразования», «удельная энергоемкость» и «потенциальная удельная энергоспособность» как ключевыми символами в «Идее единого гомеостатического устройства». Более подробно эти понятия будут рассмотрены нами в части второй: «Участники гомеостатического мира (Системный анализ частного)».
[36] от латинского informatio
[37] В своих оригинальных работах формульным символом вариантов форм, участвующих в многообразии, использован символ H. Мы невольно вынуждены были заменить символ H символом X в транскрипциях формул, чтобы не путать себя и читателя последующим изложением, где символ H будет применен, как общепринятое обозначение энтальпии системы.
[38] Рассмотрев только один частный случай нормального распределения, мы имели целью не подробные выкладки математической статистики, а лишь конкретную иллюстрацию принципиальных отношений информации (I) и вероятности события (X), распространимых, при необходимости, на другие случайные события и их вероятности.
[39] Очень интересно, и еще, наверное, более важно, следующее. Потрясшее когда-то мир заявление древнегреческого философа Сократа (469-399 до н.э.): «Единственное, что я знаю, так это то, что я ничего не знаю». И яркие принципы его ученика и последователя Платона (428/426-348/347 до н.э.), блистательно представленные в знаменитых «Платоновых кругах знания» и комментариях к ним: «чем больше круг знаний, тем больше поверхность его соприкосновения с незнанием». И теорема «неполноты формальных систем» австрийского логика и математика Курта Геделя (1906-1978), доказывавшей «невозможность полной формализации знаний». По своей коренной сути, от древнего до современного, они все содержали тот ответ, который мы только что сформулировали, говоря, что есть, и как вычислить количество информации, заставить его обрести свое строгое математическое выражение, имеющее очевидный физический смысл.
Оглавление Введение Часть 1 Часть 2 Часть 3 Заключение
Глава 1 Глава 2 Глава 3 Глава 4 Глава 5 Литература